

Translate this page into:
Biochar and vermicompost modulated Pb toxicity in summer savory (Satureja Hortensis L.) plants through inducing physiological and biochemical changes
⁎Corresponding author. hakimi.leila@iau.ac.ir (Leila Hakimi)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
The primary environmental factor affecting the biochemical attributes of plants is soil pollution by heavy metals. The detrimental effects of heavy metals on plants can be lessened by soil amendments. The goal of the current research was to determine the potential of biochar (BC) and vermicompost (VC) as soil amendments in reducing Pb toxicity in summer savory (Satureja Hortensis L.) though physiological and biochemical modifications. Therefore, the pot experiment was conducted with Pb toxicity (control (non-Pb), 300, and 600 mg kg soil−1) and soil amendments (control, 2 % BC, 10 % VC, and 1 % BC + 5 % VC) based on a completely randomized design. The results showed that Pb toxicity at 600 mg kg soil−1 led to significant decreases in shoot weight (41 %), root weight (25 %), leaf relative water content (20 %), and chlorophyll content (39 %) compared to the control. However, it resulted in increases in malondialdehyde (61 %) and electrolyte leakage (49 %) when BC and VC were not applied. The results showed that Pb toxicity at 600 mg kg soil−1 led to decreases in shoot weight (41 %), root weight (25 %), leaf relative water content (20 %), and chlorophyll content (39 %), but increases in malondialdehyde (61 %), and electrolyte leakage (49 %) compared to the control in the treatments without BC and VC application. However, BC and VC, particularly their combination were more effective in improving plant growth. The interaction of Pb at 300 mg kg−1 and combined BC and VC resulted in higher total phenolic content, total flavonoid content, essential oil (EO) content, and EO yield with 29, 62, and 39, and 35 % raises compared with the control. Agglomerative hierarchical clustering revealed that Pb at 300 and 600 mg kg soil−1 differed from the control, and that combined VC and BC significantly varied from their individual values. Combining BC and VC is more effective than using them separately in alleviating Pb toxicity, as it enhances plant growth and secondary metabolite production. The results have the potential to benefit the improvement of summer savory resistance in Pb-polluted soils.
Keywords
Heavy metal toxicity
Organic fertilizer
Biochemistry
Essential oil

1 Introduction
Heavy metals are defined as metals with an atomic number greater than 20 and a density larger than 3 g cm−3. Some of these metals, such as Cu, Zn, Ni, Mn, and Fe, are low-use elements that are crucial for natural growth, oxidation–reduction reactions, electron transfer, and participation in a variety of other metabolic processes, but their excess amount in the soil causes metabolic disorders and growth inhibition in most plant species (Khosropour et al., 2019; Mohan et al., 2021). Many heavy metals, including Pb, Cd, Cr, and Hg, are not required and toxic to plants even at low concentrations. The presence of heavy metals in the environment of plants is a stress factor that causes physiological changes in plants and can reduce the plant's growth and even cell death under a severe stress (Khosropour et al., 2019; Rahman et al., 2019). Lead is a toxic metal for humans and a non-essential metal for plants with unknown biological functions, but because it is soluble in water—in the form of lead nitrate, which has a solubility of 3.54 g/100 g of water at room temperature—it is easily absorbed by plant roots and has an impact on their growth and metabolism. Lead toxicity reduces growth and yield, yellowing of young leaves, absorption of some essential elements like iron, and photosynthesis (Zulfiqar et al., 2019). This is because Pb prevents the absorption of minerals like Fe and manganese by blocking the entry of or forming a link with the transporters of these elements, which ultimately prevents its absorption by the roots and leaves and affects the growth of the plant (Zulfiqar et al., 2019; Sharma et al., 2022). According to National Pharmacopoeia Commission (2020), the permissible concentration of Pb element in plants is 5 mg/kg. The highest amount of Pb is absorbed by plants through roots and leaves. Plant poisoning, a reduction in cell expansion, a reduction in cell division, and a decrease in transpiration are all effects of these conditions (Zulfiqar et al., 2019; Sharma et al., 2022).
To increase the quantity and quality of food produced per unit area by integrating mineral nutrition techniques and plant organic matter, it was thought that the use of organic processes in the nutrition of medicinal plants would be a fundamental solution for the development of integrated plant nutrition management systems (Khosropour et al., 2023). In sustainable agricultural systems, it is essential to use renewable resources that have maximum ecological benefits and minimum environmental damage. One of the methods of increasing plants' resistance to stress is the use of vermicompost (VC) and biochar (BC). Vermicompost is made by earthworms through the processing of organic wastes such as animal manure and plant residues, and it has received a lot of attention due to its significant potential for soil improvement. Vermicompost increases the water retention capacity in the soil, increases ventilation, and improves the physical and chemical properties of the soil (Raza et al., 2022). Biochar is a carbon material obtained by heating plant remains and waste in an environment containing limited or no oxygen. The thermal decomposition of biomass in an oxygen-deficient environment is called pyrolysis (Yadav et al., 2023). Biochar has high stability and is produced in order to manage waste, reduce climate change, produce energy, and improve soil properties (Khosropour et al., 2021). Biochar has affected various physical properties (such as soil structure, hydraulic conductivity, and gravity) and chemical attributes (such as acidity, cation exchange capacity, organic matter) and soil biology (microbial activity, microbial diversity, enzyme activity, microbial population) (Yadav et al., 2023). Biochar increases plant performance by providing water and minerals required for plants (Khosropour et al., 2021).
The demand for herbal products has substantially increased in an effort to support human health. Thus, medicinal plants have been widely grown on a variety of soils to fulfill human and industrial demands. Summer savory (Satureja Hortensis L.) is an annual and herbaceous plant of the Lamiaceae family, which is produced due to its high-value phenolic compounds, such as carvacrol and thymol (Memari Tabrizi et al., 2021). Summer savory is valuable as a potential source of nutrients, so it can be used to control some diseases due to its beneficial antioxidant (Ejaz et al., 2023), antifungal (Ahmadi et al., 2020), antibacterial (Farahani et al., 2023), antiviral (Bezić et al., 2009), anti-inflammatory (Uslu et al., 2003; Abdelshafeek et al., 2023), anti-diabetic (Ejaz et al., 2023), antispasmodic (Hajhashemi et al., 2000; Ejaz et al., 2023), anti-cancer, (Kennedy et al., 2018), and cytotoxic effects (Farzaneh et al., 2018).
Improvements in the use of BC and VC to enhance plant growth and yield in contaminated soils have recently attracted more attention (Di et al., 2019; Wang et al., 2021; Kheir et al., 2021; Song et al., 2022). The present study hypothesized that the combination of BC and VC can regulate plant growth under Pb toxicity through improving physiological and phytochemical attributes. According to previous works, the positive role of VC and BC in modulating heavy metals has been reported in different plant species like spinach (Zafar-ul-Hye et al., 2020), barberry (Khosropour et al., 2021), wheat (Kheir et al., 2021). However, the use of these compounds in summer savory is unknown. The research question is how much VC and BC can regulate plant growth through modulating Pb stress in summer savory. Therefore, the present study was conducted to evaluate the effect of BC and VC in modulating Pb toxicity by affecting plant growth and biochemical attributes of summer savory. The finding can be beneficial for food and pharmacological industries to grow this medicinal plant in industrial and urban areas.
2 Materials and methods
2.1 Plant materials and treatments
Summer savory seeds were purchased from the Pakan Bazr company in Iran. Soil samples were collected from an unpolluted area in an agricultural field and were sterilized in an oven. The experimental soil was a sandy loam with pH: 7.09, and EC: 0.99 dS m−1. N: 0.23 %, P: 12.1 mg kg−1; K: 255 mg kg−1, total Pb: 6.03 mg kg−1.
The factorial experiment was conducted using Pb stress and soil amendments in a completely randomized design (CRD) with five replicates. To induce metal toxicity, the soils were contaminated by lead nitrate (Pb NO3) at concentrations of 0 (control), 300, and 600 mg kg−1 soil. The soil amendments included BC and VC at four levels of 0 % (control), 2 % BC, 10 % VC, and 1 %BC + 5 %VC. The amounts of BC and VC used in each pot, with a volume of 3 L, were 60, 300, and 30 + 150 g, respectively. The experiment included 12 treatment combinations (three levels of Pb stress and four levels of soil amendments. The experiment included 12 treatment combinations (three levels of Pb stress and four levels of soil amendments). The properties of BC and VC are presented in Table 1, where BC was derived from grape tree pruning waste.
Parameter
Biochar
Vermicompost
Organic matter (%)
87.6
71.5
Organic carbon (%)
76 %
36 %
N (%)
0.44
2.65
K (%)
19
1.65
P (%)
0.35
0.52
pH
9.3
6.7
2.2 The production of biochar (BC)
The process for creating BC involved washing residues of grape trees with water, followed by an outdoor drying period. After being ground, it was poured into containers with appropriate covers and heated in the furnace for four h at a rate of 20 °C per min. Thereafter, the furnace was turned on at 500 °C and was transformed into BC at this temperature (Yuan et al., 2011).
2.3 Plant weight
The plants were completely cut from the bottom of the stem and dried in the shade. A digital scale with an accuracy of 0.001 g was used to measure the shoot weight of plants (Afshari et al., 2021).
2.4 Chlorophyll (Chl) assay
Leaf total Chl was measured using the Arnon (1949) instruction. For this, 8 mL of 80 % acetone was used to homogenize 0.2 g of fresh leaves. The mixture was centrifuged for 15 min at 4 °C. Supernatant was read using a spectrophotometer at 645 and 663 nm. The amount of total Chl was determined as follows equation (1):
2.5 Relative water content (RWC) measurement
Three procedures were taken to calculate RWC using fresh leaves. After determining the leaves' fresh weight (FW), they were submerged in distilled water for 24 h to reach their saturation weight (SW). Dry weight (DW) was then calculated for the samples after being dried in an oven until the weight remained steady as follows equation (2) (Dhopte and Manuel, 2002):
2.6 Malondialdehyde (MAD) measurement
A mixture of 0.5 g of fresh leaves and 4 mL of 20 % w/v trichloroacetic acid (TCA) with 0.5 % thiobarbituric acid was prepared. The mixture was then placed in a hot water bath at 95 °C for 25 min. After the heating step, the mixture was centrifuged at 14000 rpm for 30 min to separate the supernatant from the leaf debris. The supernatant was retained for further analysis. The retained samples were immersed in a hot water bath again for 30 min. Following this, the samples were centrifuged at 10000 rpm for 10 min. Finally, the samples were read using a 532 nm laser, in accordance with the method described by Heath and Packer (1968).
2.7 Electrolyte leakage measurement (EL)
To determine EL, fresh leaves were sampled from each pot and placed in test tubes with 10 mL of double distilled water. The tubes were closed with a plastic cap and kept at a constant temperature of 25 °C. After 12 h, the initial electrical conductivity (EC1) was measured with an EC meter (model MI306, Milwaukee WI, USA). After that, the samples were boiled at 100 °C for 20 min to release all the electrolytes. Following a 25 °C cooling of the samples, the EC2 was determined. Equation (1) was used to determine the EL (Asrar et al., 2012).
2.8 Total phenolic content (TPC)
In order to extract the TPC of the samples, leaf samples were mixed with the extraction solvent consisting of methanol (85 %) and acetic acid (15 %) in a ratio of 1:40. The extraction was obtained by shaking in the dark at 4 °C for 72 h. Then, 0.5 mL of the extracts were mixed with 2.5 mL of 10 % Folin-Ciocalteu and mixed with a vortex mixer. After 6 min, 2 mL of 7.5 % sodium carbonate was added to the mixture and kept for 1.5 h in the dark. A spectrophotometer was then used to measure the absorbance at a wavelength of 750 nm. Gallic acid (GA) was used as a standard and TPC was expressed as mg GA per g dry weight (DW) of leaf samples (Khosropour et al., 2021).
2.9 Total flavonoid content (TFC)
The quantity of TFC was determined using the aluminum chloride colorimetric method. The procedure involved combining 1.5 mL of 95 % ethanol, 0.1 mL of 10 % aluminum chloride, 0.1 mL of 1 M potassium acetate, and 2.8 mL of distilled water with 0.5 mL of the extract solution. After a 30 min storage period at room temperature, the mixture's absorbance was measured at 415 nm. A curve was created using the quercetin (QE) standard, and TFC was expressed as mg QE per g DW (Khosropour et al., 2021).
2.10 Essential oil (EO) distillation and EO yield
The plants' aerial parts were taken during the flowering stage and dried in the shade to produce the EO. The 100 g of dried samples were hydro-distilled for 3 h in a Clevenger type apparatus to determine the EO content (w/w). On the basis of the dry plant weight in each pot, the EO yield was determined (Sefidkon et al., 2006).
2.11 Data analysis
The Least Significant Difference (LSD) test was used to compare the mean values and the data at a 5 % probability level using SAS (version 9.3). Multivariate analyses were accomplished by XLSTAT.
3 Results
3.1 Plant dry weight
Pb toxicity, BC, and VC significantly (P ≤ 0.05) modified the plant weight. Under non-application of VC and BC, Pb at 300 and 600 mg Kg−1 soil decreased shoot weight by 12 and 41 % (Fig. 1a) and root weight by 13 and 25 % (Fig. 1b), respectively. The BC, VC, and BC + VC respectively led to 12, 15, and 20 % declines in shoot weight (Fig. 1a) and 14, 16, and 25 % decreases in root weight Fig. 1b) in summer savory plants exposed to 600 mg Pb Kg−1 soil. In addition, under 300 mg Pb Pb Kg−1 soil, the use VC + BC led to 12 and 10 % increases in shoot and root weight, respectively. Therefore, the combination of VC and BC is eminent in modulating Pb toxicity by improving plant weight.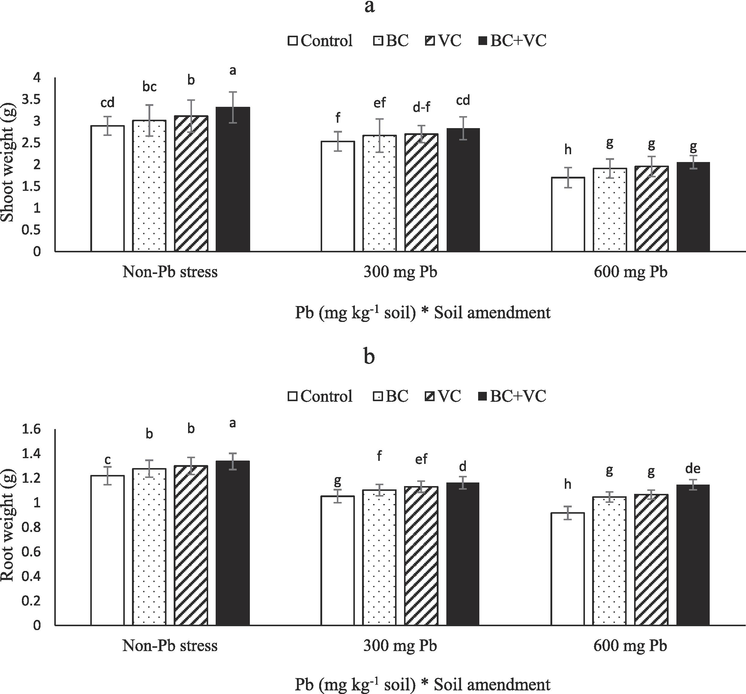
Shoot weight (a) and root weight (b) of summer savory plants under soil amendments (biochar and vermicompost) in Pb-polluted soils. Values are means ± standard deviation (SD). Different letters represent statistically significant differences among treatments at P ≤ 0.05.
3.2 Relative water content (RWC) and chlorophyll (Chl) content
The RWC and Chl were significantly reduced by Pb toxicity but increased by BC and VC. Without soil amendments, 8 and 20 % reductions in RWC were obtained for plants exposed to 300 and 600 mg Pb Kg−1 soil, respectively, compared to the control. However, the combination of BC and VC increased plant RWC by 12 % compared to the control (Fig. 2a). Like RWC, Chl showed a reduction under Pb stress but it improved by BC and VC. The maximum Chl a + b was obtained in plants with BC + VC without Pb toxicity. However, the minimum Chl was reported in the plants exposed to 600 mg Pb Kg−1 soil without BC or VC, with a 29 % reduction compared with the control (Fig. 2b).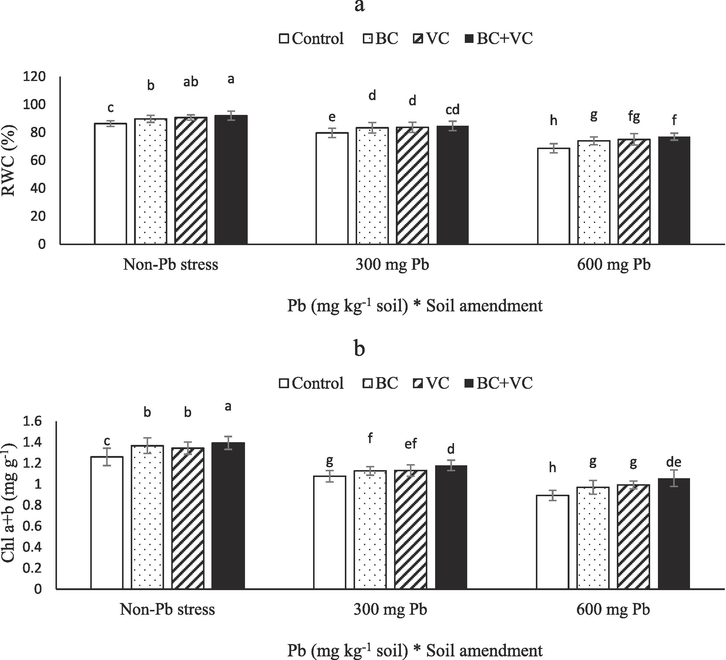
Relative water content (RWC, a) and chlorophyll (Chl) a + b (b) of summer savory plants under soil amendments (biochar and vermicompost) in Pb-polluted soils. Values are means ± standard deviation (SD). Different letters represent statistically significant differences among treatments at P ≤ 0.05.
3.3 Malondialdehyde (MDA) and electrolyte leakage (EL)
The MDA significantly increased by Pb toxicity, while BC and VC lowered it. The maximum MDA was observed in plants exposed to 600 mg Pb Kg−1 soil, with a 61 % enhancement compared with the control. The BC, VC, and BC + VC lowered MDA by 14, 19, and 23 % in plants under 600 mg Pb Kg−1 soil (Fig. 3a). In addition, the results revealed 27 and 49 % raises of EL when plants experienced 300 and 600 mg Pb Kg−1 soil, respectively. The best results of soil amendments were related to the combined application of BC and VC by a 15 % decline in EL compared with non-BC/VC application in plants under severe Pb stress (Fig. 3b).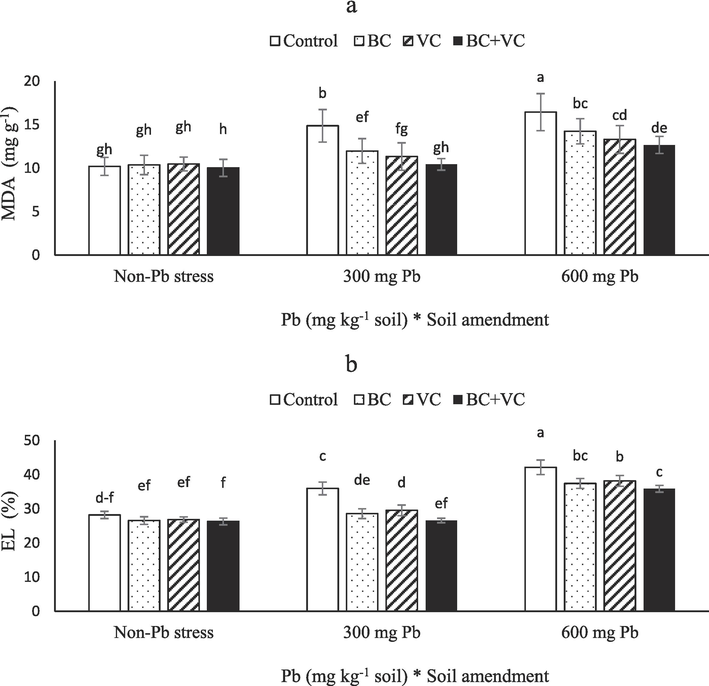
Malondialdehyde (MDA, a) and electrolyte leakage (EL, b) of summer savory plants under soil amendments (biochar and vermicompost) in Pb-polluted soils. Values are means ± standard deviation (SD). Different letters represent statistically significant differences among treatments at P ≤ 0.05.
3.4 Total phenolic content (TPC) and total flavonoid content (TFC)
Higher TPC was triggered by Pb exposure, and also BC and VC either alone or together improved TPC. Upon VC + BC, TPC increased 14 and 17 % after the application of 300 and 600 mg Pb Kg−1 soil, respectively. compared with control, a 32 % enhancement in TFC was observed in the interaction of 600 mg Pb Kg−1 soil and BC + VC, (Fig. 4a). TFC increased as a result of moderate Pb toxicity and VC, reaching a maximum in plants treated with 300 mg Pb Kg−1 soil and VC, a 67 % improvement over control (Fig. 4b).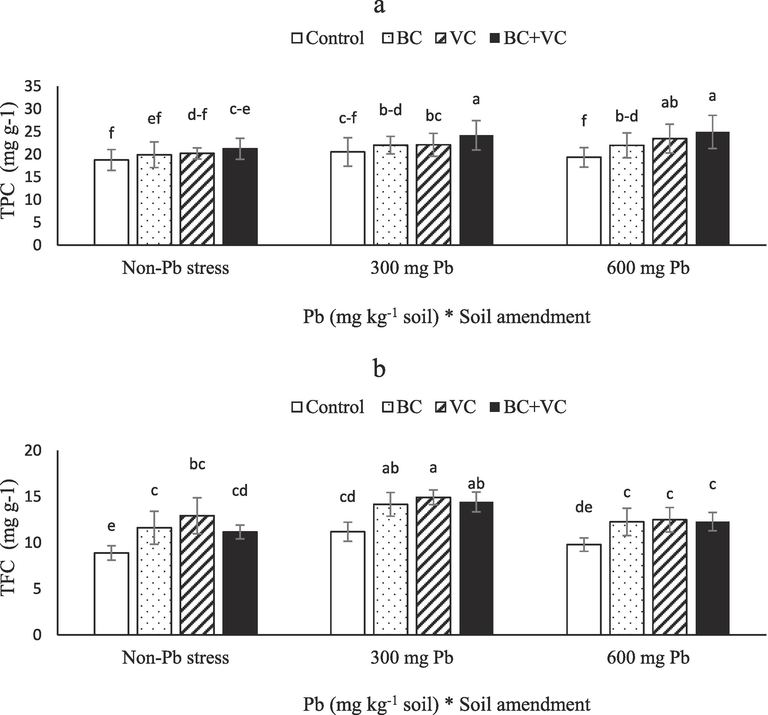
Total phenolic content (TPC, a) and total flavonoid content (TFC, b) of summer savory plants under soil amendments (biochar and vermicompost) in Pb-polluted soils. Values are means ± standard deviation (SD). Different letters represent statistically significant differences among treatments at P ≤ 0.05.
3.5 Essential oil (EO) content and EO yield
Interesting results were obtained for EO content with an increasing trend at 300 mg Pb Kg−1 soil. The EO content in the interaction of 300 mg Pb Kg−1 soil and BC + VC was higher than other treatments, with 38 % enhancement relative to the control (Fig. 5a). Although EO yield in treatments containing 300 mg Pb Kg−1 soil had no significant difference with the non-Pb stress, Pb toxicity at 600 mg Kg−1 soil led to noticeable declines. Accordingly, the minimum EO yield was reported in the 600 mg Pb Kg−1 soil without BC/VC with 35 % decreases relative to the control (Fig. 5b).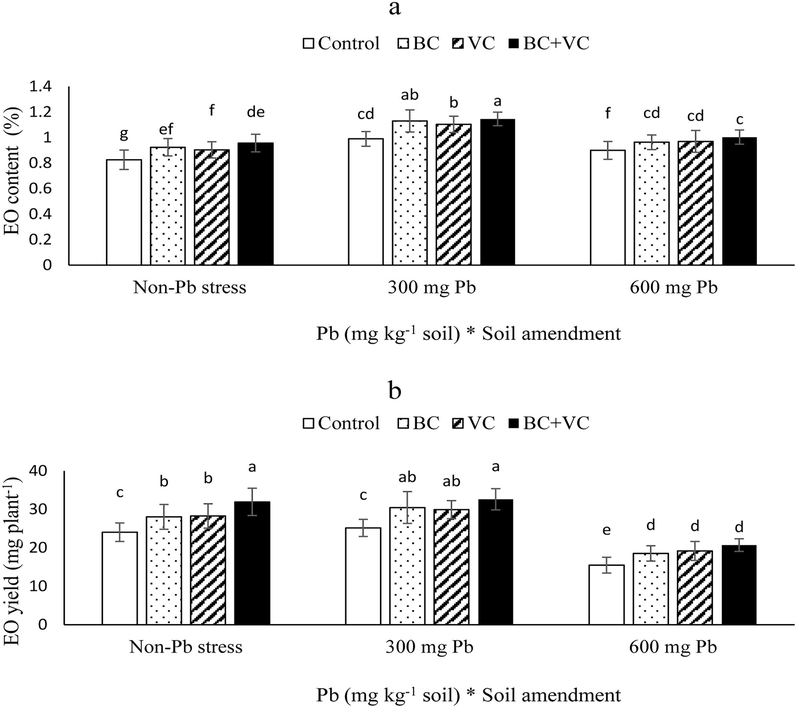
Essential oil (EO) content (a) and EO yield (b) of summer savory plants under soil amendments (biochar and vermicompost) in Pb-polluted soils. Values are means ± standard deviation (SD). Different letters represent statistically significant differences among treatments at P ≤ 0.05.
3.6 Multivariate analysis
According to the principal component analysis (PCA) results for Pb toxicity, F1 (axis 1) explained control and 600 mg Pb Kg−1 soil, whereas F2 described 300 mg Pb Kg−1 soil. Additionally, all traits were supported by F1 except for EO content and TFC (Fig. 6a). However, soil amendment PCA revealed that most traits were supported by F1. VC, BC + VC, and control were described by F1 while BC was defended by F2 (Fig. 6b). Additionally, shoot weight, root weight, RWC, Chl, and EO yield negatively correlated with MDA, EL, and TPC.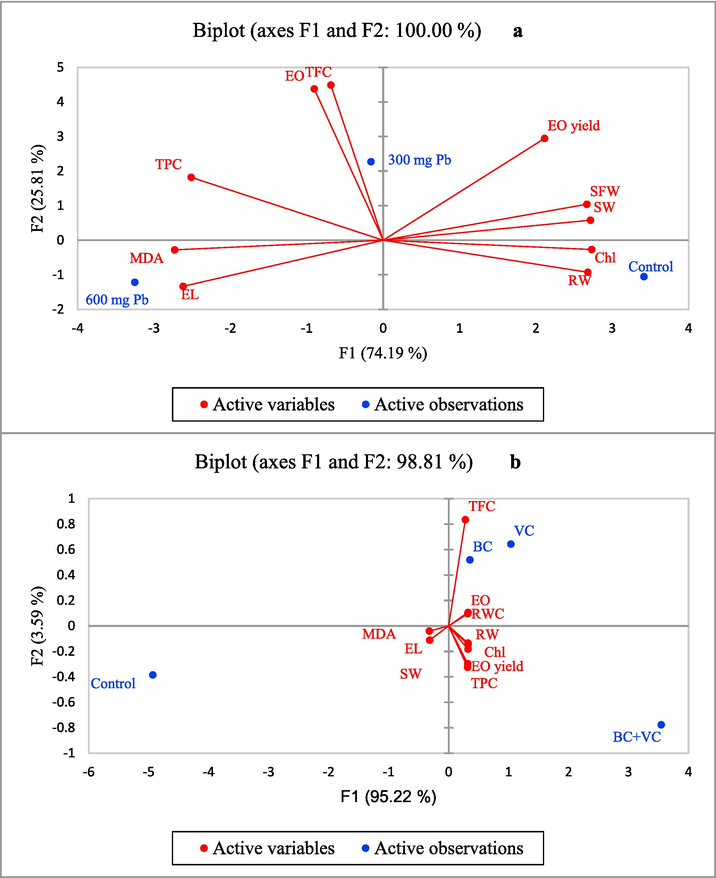
Principal component analysis (PCA) of the traits under Pb stress (a) and soil amendments (b). BC: biochar, VC: vermicompost, SW: shoot weight, RW: root weight, MDA: malondialdehyde, EL: electrolyte leakage, EO: essential oil, EOY: essential oil yield, RWC: relative water content. TPC: total phenolic content, TFC: total flavonoid content.
Three distinct clusters were identified based on agglomerative hierarchical clustering (AHC) of Pb toxicity as each level was placed in a cluster (Fig. 7a). Additionally, three clusters were identified for soil amendments as BC and VC in one cluster, BC + VC in another cluster, and control in yet another cluster (Fig. 7b).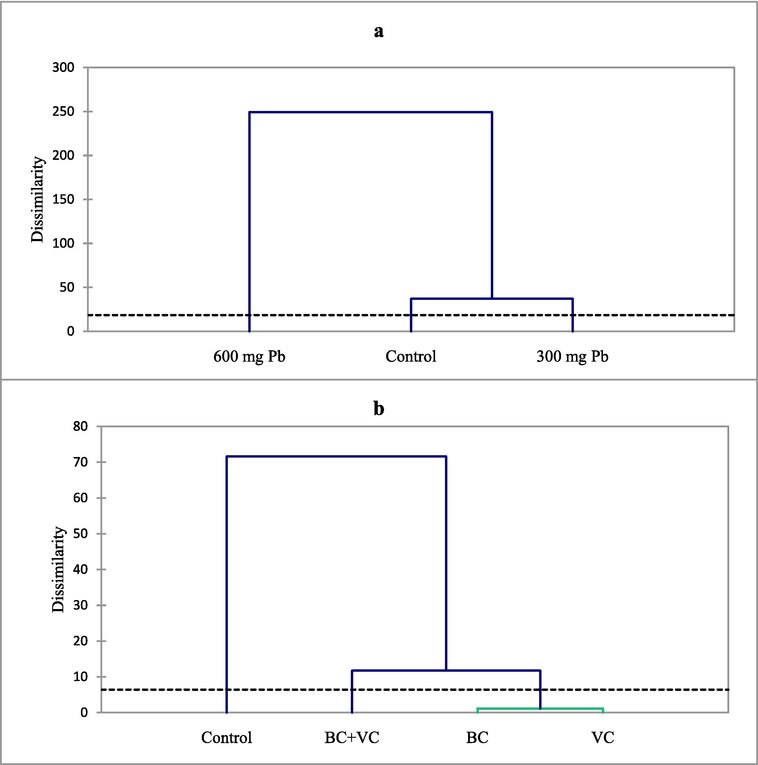
Agglomerative hierarchical clustering (AHC) of the traits under Pb stress (a) and soil amendments (b).
4 Discussion
Lead stress severely limited plant growth, although VC and BC significantly enhanced it. It appears that heavy metals like Pb compete with other nutrients like calcium (Ca), iron (Fe), manganese (Mn), copper (Cu), nickel (Ni), zinc (Zn), potassium (K), and sodium (Na) for absorption by membrane transporters, which can result in a deficiency of these vital nutrients in the plant and slow down plant growth (Irfan et al., 2021). Lead interferes with the availability of soil minerals and hinders plants' ability to absorb nutrients, which limits their growth. Previous studies have revealed that Pb plays an important role in the inhibition of nutrient movement to the leaves (Zafar-ul-Hye et al., 2020). Leaf induces excessive production of ROS when the plant's defense network is not able to maintain free concentrations of Pb ions in the cytosol at a level lower than the threshold. Also, Pb changes the activity of key enzymes of many metabolic pathways. These changes may be the result of inhibiting some primary physiological processes such as nitrogen fixation and other metabolic processes (Zafar-ul-Hye et al., 2020). The retention of Pb in the root may also cause a further reduction in root growth as compared to the aerial parts. Additionally, Pb toxicity limits water transfer to growing tissues and leaves, alters the ultrastructure of cellular organelles, and alters the activity of numerous enzymes involved in metabolic pathways. These effects have a negative impact on seed germination and plant growth (Zulfiqar et al., 2019). Higher accumulation of Pb in plants may interfere with plant growth and development in various ways, such as reducing enzyme activities, disrupting photosynthesis and dark respiration, closing stomata, and inhibiting water absorption. Lead prevents the division of cells in the meristem region and the growth of cells in the growth region. On the other hand, early differentiation and lignification of the cell wall located in the longitudinal growth region of the cell can be one of the reasons for the reduction of root growth (Ivanov and Zhukovskaya, 2021). Mainly, lead enters the root through an apoplastic pathway or Ca ion channel. The transfer of lead from the apoplastic pathway is easily done through the dissolution of Pb in water (Kumar et al., 2020). This study supports the beneficial role of BC and VC as a useful technique to reduce Pb accumulation in summer savory plants. Organic substrates help plants tolerate Pb stress by decreasing Pb bioavailability and mobility in the soil, limiting Pb absorption and translocation, storing Pb in the vacuole, and controlling transpiration rate. The BC can enhance the soil's chemical characteristics, including its pH and electrical conductivity (EC). Biochar is filling the pH gap between the soil and this organic material through soil pH enhancement (Albert et al., 2021). Like our results, the beneficial role of BC and VC in modulating heavy metal toxicity has been reported on barberry (Khosropour et al., 2021) and wheat (Kheir et al., 2021). Overall, the combination of VC and BC offers a comprehensive approach to improving soil fertility, structure, and plant growth. Their synergistic effects in nutrient enrichment, immobilization of Pb, and promotion of beneficial microbial activity make them a more effective combination than their individual applications (Wang et al., 2018).
Lead toxicity reduced RWC content, while it was improved by VC and BC. The RWC is a significant factor in plants under stress conditions because it monitors changes in plant water status through stomatal conductance (Ma et al., 2023). Pb toxicity in plants reduces RWC through several biochemical mechanisms. It increases oxidative stress, leading to membrane lipid peroxidation and cellular damage that impairs water movement and reduces water uptake. Pb toxicity also disrupts the functioning of aquaporins, affecting water transport across cell membranes. It can alter stomatal behavior, causing stomatal closure or malfunction, and reducing transpiration and water loss control (Bakhtiari et al., 2023). Furthermore, Pb toxicity disturbs root hydraulic conductivity, hindering water uptake and flow from roots to shoots. These biochemical pathways collectively contribute to the decrease in RWC in plants exposed to Pb toxicity (Ma et al., 2023; Bakhtiari et al., 2023). The application of biochar and vermicompost can improve RWC in plants under Pb toxicity through various mechanisms, including enhanced water holding capacity, stimulation of root growth, regulation of stomatal behavior, induction of osmotic adjustment, and activation of stress-responsive genes and proteins. These mechanisms contribute to maintaining proper water balance, optimizing water uptake, and reducing dehydration effects caused by Pb toxicity (Kheir et al., 2021). It has been reported that when VC and BC were used in combination, the soil physical and chemical attributes were much better than their separate use (Huang and He, 2023). Similarly, Khosropour et al. (2021) showed that the combined VC and BC was more effective than their separate uses in improving RWC in barberry plants exposed to Cd stress.
Lead had a negative impact on Chl content, but BC and VC improved it. Inhibition of important enzymes such as delta-aminolevulinic acid dehydratase, which are involved in chlorophyll biosynthesis, is also one of the possible reasons for the reduction of chlorophyll under Pb toxicity. Photosynthesis is very sensitive to the disruption of the movement of gases from the stomata, and electron microscope observations show that the stomata are closed by Pb. Reactive oxygen species are the inevitable consequence of plant response to Pb stress. The decrease in photosynthesis rate during Pb stress can be attributed to stomatal factors (Ramyar et al., 2022). Stomatal limitations of photosynthesis can be described as inefficient absorption of CO2 in leaves and sub-stomatal cavities due to premature closure of stomata. Reduced Rubisco enzyme activity, chloroplast activity, ATP availability, ribulose 1,5-bs phosphate synthesis, leaf nitrogen, and disruption of photosystems I and II are all factors that limit non-photosynthetic photosynthesis (Ramyar et al., 2022). The reduction of Chl content under Pb stress is considered a typical sign of pigment photooxidation and Chl degradation. The combined effect of BC and VC significantly acts as a nutritional source as well as a suitable cation exchanger due to its large surface area and as a result of the availability of nutrients and their greater absorption by plant roots. In addition, the increase in soil pH amended by BC has consequences on mineralization and availability of nutrients, mainly related to basic cations such as Ca, Mg, and K (Huang and He, 2023).
The MDA and EL increased by Pb toxicity, but decreased by BC and VC. Cell membrane markers like MDA and EL play a major role in predicting plant stress. Increased MDA and EL are indicators of plant damage and are primarily caused by elevated ROS in stressful situations (Okant and Kaya et al., 2019). Zeng et al. (2021) represented that Pb toxicity at 300 and 600 mg Kg−1 significantly increased MDA and EL in Castorbean, which is consistence with the present findings. Soil amendments have a positive effect on antioxidant capacity, resulting in lower MDA and EL. The modulation role of BC and VC for Pb toxicity can be detected as its effect on cell and membrane stability. The reduced MDA and EL by BC and VC have been discovered in eggplant by Ebrahimi et al. (2021).
The interaction of Pb at 300 mg Kg−1 and combined BC and VC led to higher TPC and TFC. The phenylpropanoid compounds of plants cause color, taste, and special physiological characteristics in plants and protect the plant against biotic and abiotic stresses (Cheynier et al., 2013). Phenolic compounds are related to their oxidation–reduction properties, which play an important role in absorbing and neutralizing free radicals, suppressing ROS, or decomposing peroxidases (Khosropour et al., 2021). In most plant species, the key step in making phenolic compounds is converting phenylalanine to cinnamic acid and it is done with the help of phenylalanine ammonialyase (PAL) enzyme. The present work showed that moderate Pb toxicity with combined BC and VC can be effective in reaching the maximum TPC and TFC. Similar to the results, the highest TPC and TFC have been reported in coriander plants by Babashpour-Asl et al. (2022) under moderate Cd toxicity and Afshari et al. (2021) under mild water stress. The soil amendments often include special compounds that activate the plant's defense strategy by inducing false stress and as a result produce secondary metabolites. Many studies show that the addition of VC and BC can modulate different abiotic stresses by improving the water and nutritional attributes. The production pathway of phenolic compounds can be induced by BC and VC due to the changes in nutritional value in plants. In addition, BC and VC can change the transcription of genes and enzymes involved in the production of phenolic compounds. Khosropour et al. (2021) represented that the maximum TPC and TFC in barberry plants were obtained with moderate Cd stress (10 mg kg−1 soil) with combined BC and VC, which are in line with our results.
EO content and EO yield decreased by severe Pb toxicity. However, the interaction between moderate Pb stress and the application of BC and VC can potentially enhance the EO content in plants up to a maximum level. BC and VC can enhance soil fertility and nutrient availability, promoting optimal plant growth and nutrient uptake. EO biosynthesis is tightly linked to nutrient availability, particularly nitrogen, phosphorus, and potassium. The application of BC and VC can provide an abundant supply of these nutrients, thus supporting the synthesis and accumulation of essential oils in plants (Sharafabad et al., 2022). Moderate heavy metal toxicity has been reported to stimulate EOs in various plant species, while their severe toxicity led to decreased EO content (Memari-Tabrizi et al., 2021; Babashpour-Asl et al., 2022; Bakhtiari et al., 2023, Alawamleh et al., 2023). Co-application of BC and VC and Pb toxicity at 300 mg kg soil−1 (moderate stress) can enhance the activity of enzymes involved in EO biosynthesis, such as terpene synthases, phenylpropanoid enzymes, and antioxidants (Memari-Tabrizi et al., 2021). This stimulation of secondary metabolite production can lead to increased EO content in plants under moderate Pb stress and soil-applied BC and VC.
Changes in plant weight have an obvious effect on EO yield since EO yield is estimated using plant weight and EO content. Plants exposed to metal toxicity slow down their secondary metabolism, which affects the metabolic processes that produce EO. The abiotic stress led to changes in the number and size of EO glands in leaves as the high rate of stress led to decreased size and number of glands (Gohari et al., 2020). Memari-Tabrizi et al. (2021) on summer savory plants and Babashpour-Asl et al. (2022) on coriander plants addressed the declines in EO content and yield under 20 mg Cd kg−1 soil.
All phases of plant life, from seed germination to plant development, can be impacted by heavy metals like Pb. It has been reported that auxin and abscisic acid, two main phytohormones, are altered by Pb during seed germination. Meristematic cells' ability to divide mitotically is inhibited, which results in a decrease in root length and weight and an increase in root diameter. Lead also alters the relationship between nutrients and water for plants, which has an impact on the production of secondary metabolites like EO. It restricts mitotic division of meristematic cells, which results in a decrease in root length and weight and an increase in root width. Lead also alters the relationship between nutrients and water for plants, which has an impact on the production of secondary metabolites like EO (Lermen et al., 2015). However, when summer savory plants were exposed to Pb toxicity, the combined Bc and VC increased EO yield. Specific functions of BC and VC in plants include improved water absorption, nutrient uptake, gas exchange, and phytohormone adjustment (Khosropour et al., 2021). Improvement of EO yield with BC and VC has been addressed on garlic (Ghassemi and Raei, 2021) and Cymbopogon martini (Roxb.) Wats (Jain et al., 2020).
5 Conclusions
The current findings demonstrated that severe lead toxicity considerably reduced plant development and the main secondary metabolites, but that biochar and vermicompost, especially their combination, effectively controlled lead toxicity by increasing chlorophyll and water content. In order to reduce lead toxicity in summer savory plants, it is suggested to employ a combination of biochar and vermicompost, as multivariate research revealed that this was significantly different from using either material separately. The lead stress at 300 mg Kg−1 can be advised to get the maximal secondary metabolites, such as essential oil content and phenolic compounds. The results of the current study demonstrated that using vermicompost and biochar together can effectively reduce the effects of heavy metal stress on medicinal plants. This finding merits future investigations on medicinal plants.
Author contribution
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by all authors. The first draft of the manuscript was written by Ayda Alavian and revised by Noushin Osuleddini and Leila Hakimi. All authors read and approved the final manuscript.
Declaration of competing interest
The author declares the following financial interests/personal relationships which may be considered as potential competing interests: Leila Hakimi reports was provided by AL-Balqa Applied University. Leila Hakimi reports a relationship with Islamic Azad University as a researching membership.
References
- Phytochemical profile, comparative evaluation of Satureja montana alcoholic extract for antioxidants, anti-inflammatory and molecular docking studies. BMC Complement. Med. Therap.. 2023;23:1-12.
- [CrossRef] [Google Scholar]
- Foliar-applied silicon and its nanoparticles stimulate physio-chemical changes to improve growth, yield and active constituents of coriander (Coriandrum Sativum L.) essential oil under different irrigation regimes. Silicon. 2021;13:4177-4188.
- [CrossRef] [Google Scholar]
- Anti-bacterial/fungal and anti-cancer performance of green synthesized Ag nanoparticles using summer savory extract. J. Exp. Nanosci.. 2020;15:363-380.
- [CrossRef] [Google Scholar]
- Cold plasma and foliar-applied selenium nanoparticles modulated cadmium toxicity through changes in physio-biochemical properties and essential oil profile of sage (Salvia officinalis L.) J. Soil Sci. Plant Nutr. 2023:1-15.
- [CrossRef] [Google Scholar]
- Influence of biochar and soil properties on soil and plant tissue concentrations of Cd and Pb: A meta-analysis. Sci. Total Environ.. 2021;755:142582
- [CrossRef] [Google Scholar]
- Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol.. 1949;24:1-15.
- [Google Scholar]
- Improving growth, flower yield, and water relations of snapdragon (Antirhinum majus L.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi. Photosynthetica. 2012;50:305-316.
- [CrossRef] [Google Scholar]
- Foliar-applied selenium nanoparticles alleviate cadmium stress through changes in physio-biochemical status and essential oil profile of coriander (Coriandrum sativum L.) leaves. Environ. Sci. Pollut. Res.. 2022;29:80021-80031.
- [CrossRef] [Google Scholar]
- Foliar application of silicon, selenium, and zinc nanoparticles can modulate lead and cadmium toxicity in sage (Salvia officinalis L.) plants by optimizing growth and biochemical status. Environ. Sci. Pollut. Res.. 2023;30:54223-54233.
- [CrossRef] [Google Scholar]
- Essential oil composition and internal transcribed spacer (ITS) sequence variability of four South-Croatian Satureja species (Lamiaceae) Molecules. 2009;14:925-938.
- [CrossRef] [Google Scholar]
- Plant phenolics: recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem.. 2013;72:1-20.
- [CrossRef] [Google Scholar]
- Dhopte, A.M., Manuel, L.M., 2002. Principles and Techniques for Plant Scientists, 1st end. Updesh Purohit for Agrobios (India), Odhpur, ISBN, 81, 373.
- Biochar combined with vermicompost increases crop production while reducing ammonia and nitrous oxide emissions from a paddy soil. Pedosphere. 2019;29:82-94.
- [CrossRef] [Google Scholar]
- Biochar and vermicompost improve growth and physiological traits of eggplant (Solanum melongena L.) under deficit irrigation. Chem. Biol. Technol. Agric.. 2021;8:1-14.
- [CrossRef] [Google Scholar]
- A comprehensive review of summer savory (Satureja hortensis L.): promising ingredient for production of functional foods. Front. Pharmacol.. 2023;14
- [CrossRef] [Google Scholar]
- Antimicrobial and antioxidant effects of an edible coating of Lepidium sativum seed mucilage and Satureja hortensis L. essential oil in uncooked lamb meat. Food Control 2023110240
- [CrossRef] [Google Scholar]
- Inhibition of LPS-induced inflammatory responses by Satureja hortensis extracts in J774. 1 macrophages. J. Immunoass. Immunoch.. 2018;39:274-291.
- [CrossRef] [Google Scholar]
- Evaluation of ion content, productivity and essential oil quality of garlic under saline conditions and biochar and polyamine treatments. J. Food Compos. Anal.. 2021;96:103720
- [CrossRef] [Google Scholar]
- Titanium dioxide nanoparticles (TiO2 NPs) promote growth and ameliorate salinity stress effects on essential oil profile and biochemical attributes of Dracocephalum moldavica. Sci. Rep.. 2020;10:1-14.
- [CrossRef] [Google Scholar]
- Antispasmodic and anti-diarrhoeal effect of Satureja hortensis L. essential oil. J. Ethnopharmacol.. 2000;71:187-192.
- [CrossRef] [Google Scholar]
- Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys.. 1968;125:189-198.
- [CrossRef] [Google Scholar]
- Impacts of biochar and vermicompost addition on physicochemical characteristics, metal availability, and microbial communities in soil contaminated with potentially toxic elements. Sustainability. 2023;15:790.
- [CrossRef] [Google Scholar]
- Heavy metals immobilization and improvement in maize (Zea mays L.) growth amended with biochar and compost. Sci. Rep.. 2021;11:1-9.
- [CrossRef] [Google Scholar]
- Effect of heavy metals on root growth and the use of roots as test objects. Russian J. Plant Physiol.. 2021;68:S1-S25.
- [CrossRef] [Google Scholar]
- Biochar aided aromatic grass [Cymbopogon martini (Roxb.) Wats.] vegetation: A sustainable method for stabilization of highly acidic mine waste. J. Hazard. Mat.. 2020;390:121799
- [CrossRef] [Google Scholar]
- Lung cancer stage-shift following a symptom awareness campaign. Thorax. 2018;73:1128-1136.
- [CrossRef] [Google Scholar]
- Biochar blended humate and vermicompost enhanced immobilization of heavy metals, improved wheat productivity, and minimized human health risks in different contaminated environments. J. Environ. Chem. Engineer.. 2021;9:105700
- [CrossRef] [Google Scholar]
- Response of Platanus orientalis leaves to urban pollution by heavy metals. J. For. Res.. 2019;30:1437-1445.
- [CrossRef] [Google Scholar]
- Vermicompost and biochar can alleviate cadmium stress through minimizing its uptake and optimizing biochemical properties in Berberis integerrima bunge. Environ. Sci. Pollut. Res.. 2021;1–11
- [CrossRef] [Google Scholar]
- Role of engineered nanomaterials in biotic stress managements. In: Engineered Nanomaterials for Sustainable Agricultural Production, Soil Improvement and Stress Management. Academic Press; 2023. p. :257-272.
- [CrossRef] [Google Scholar]
- Kumar, A., Kumar, A., M.M.S., C.P., Chaturvedi, A.K., Shabnam, A.A., Subrahmanyam, G. Yadav, K.K., 2020. Lead toxicity: health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health. 17, 2179. Doi: 10.3390/ijerph17072179.
- Essential oil content and chemical composition of Cymbopogon citratus inoculated with arbuscular mycorrhizal fungi under different levels of lead. Ind. Crop Prod.. 2015;76:734-738.
- [CrossRef] [Google Scholar]
- Chemical and mechanical coating of sulfur on baby corn biochar and their role in soil Pb availability, uptake, and growth of tomato under Pb contamination. Environ. Pollut.. 2023;338:122654
- [CrossRef] [Google Scholar]
- Foliar-applied silicon nanoparticles mitigate cadmium stress through physio-chemical changes to improve growth, antioxidant capacity, and essential oil profile of summer savory (Satureja hortensis L.) Plant Physiol. Biochem.. 2021;165:71-79.
- [CrossRef] [Google Scholar]
- Phytoremediation of heavy metals from the biosphere perspective and solutions. Pollut. Water Manage. 2021:95-127.
- [CrossRef] [Google Scholar]
- The role of endogenous nitric oxide in melatonin-improved tolerance to lead toxicity in maize plants. Environ. Sci. Pollut. Res.. 2019;26:11864-11874.
- [CrossRef] [Google Scholar]
- The relative impact of toxic heavy metals (THMs) (arsenic (As), cadmium (Cd), chromium (Cr)(VI), mercury (Hg), and lead (Pb)) on the total environment: an overview. Environ. Monitor. Assess.. 2019;19:1-21.
- [CrossRef] [Google Scholar]
- Reduction of lead toxicity effects and enhancing the glutathione reservoir in green beans through spraying sulfur and serine and glutamine amino acids. Environ. Sci. Pollut. Res.. 2022;1–17
- [CrossRef] [Google Scholar]
- Reuse of agricultural wastes, manure, and biochar as an organic amendment: A review on its implications for vermicomposting technology. J. Clean. Prod. 2022132200
- [CrossRef] [Google Scholar]
- Influence of drying and extraction methods on yield and chemical composition of the essential oil of Satureja hortensis. Food Chem.. 2006;99:19-23.
- [CrossRef] [Google Scholar]
- Integrated humic acid and vermicomposting changes essential oil quantity, and quality in field-grown Lavandula angustifolia L. intercropped with Brassica nigra L. Ind. Crop. Prod.. 2022;178:114635
- [CrossRef] [Google Scholar]
- Heavy metal induced regulation of plant biology: Recent insights. Physiol. Plant.. 2022;174:e13688.
- [Google Scholar]
- Biochar/vermicompost promotes Hybrid Pennisetum plant growth and soil enzyme activity in saline soils. Plant Physiol. Biochem.. 2022;183:96-110.
- [CrossRef] [Google Scholar]
- Effects of aqueous extracts of Satureja hortensis L. on rhinosinusitis treatment in rabbit. J. Ethnopharmacol.. 2003;88:225-228.
- [CrossRef] [Google Scholar]
- Biochar and vermicompost improve the soil properties and the yield and quality of cucumber (Cucumis sativus L.) grown in plastic shed soil continuously cropped for different years. Agric. Ecosyst. Environ.. 2021;315:107425
- [CrossRef] [Google Scholar]
- Vermicompost and biochar as bio-conditioners to immobilize heavy metal and improve soil fertility on cadmium contaminated soil under acid rain stress. Sci. Total Environ.. 2018;621:1057-1065.
- [CrossRef] [Google Scholar]
- Biochar application: A sustainable approach to improve soil health. J. Agric. Food Res. 2023100498
- [CrossRef] [Google Scholar]
- The forms of alkalis in the biochar produced from crop residues at different temperatures. Biores. Technol.. 2011;102:3488-3497.
- [CrossRef] [Google Scholar]
- Potential role of compost mixed biochar with rhizobacteria in mitigating lead toxicity in spinach. Sci. Rep.. 2020;10:1-12.
- [CrossRef] [Google Scholar]
- Combined citric acid and glutathione augments lead (Pb) stress tolerance and phytoremediation of Castorbean through antioxidant machinery and Pb uptake. Sustainability. 2021;13:4073.
- [CrossRef] [Google Scholar]
- Lead toxicity in plants: Impacts and remediation. J. Environ. Manag.. 2019;250:109557
- [CrossRef] [Google Scholar]




