

Translate this page into:
Date (Phoenix dactylifera) seed and syringic acid exhibits antioxidative effect and lifespan extending properties in Caenorhabditis elegans
⁎Corresponding author. birendra-bot@patnauniversity.ac.in (Birendra Prasad)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Date fruit (Phoenix dactylifera (PD); Arecaceae) has numerous proven health benefits and is the staple food in the Middle East. The seed which is discarded forms one-fourth of its weight and is rich in polyphenols. However, the possible effect of PD seed (PDS) extract in Caenorhabditis elegans has not been reported. The objective of this investigation was to provide the first scientific based results of the in vivo antioxidant effect of the PDS extract and its active ingredient in a Caenorhabditis elegans animal model system. The composition of a methanol extract of PDS was analyzed by RP-HPLC. Ferric reducing antioxidant power (FRAP) and hydrogen peroxide scavenging assays were performed for the estimation of in vitro antioxidant activity of the crude extract and one of its components – syringic acid (SA). For the in vivo effect, worms were exposed to different concentrations of PDS extract and SA. Lifespan and oxidative stress response were analyzed. To investigate the pathway involved, transgenic strains (mu86, DAF-16::GFP) of C. elegans were used. The amount of syringic acid in the phenolic content of the extract was 10.72 µg/mL. PDS and SA demonstrated antioxidative property in concentration dependent fashion and a concentration of 250 µg/mL PDS and 600 µg/mL SA significantly increased the lifespan and provided indemnity against pro-oxidants. The effect was independent of the DAF-16 transcriptional regulator as observed by using DAF16::GFP nuclear translocation assays and a daf-16 null mutant strains. The fertility also increased upon treatment with PDS and SA. This implicates syringic acid as a possible drug and date seed powder can thence be further investigated to be used as a dietary nutritional supplement.
Keywords
C. elegans
Phoenix dactylifera
RP-HPLC
Syringic acid
DAF-16::GFP
Daf-16 null mutants

1 Introduction
Aging is a natural pleiotropic phenomenon inherent to all living beings. The desire to always remain youthful has driven scientists to search for novel substances that might curtail the myriad effects of ageing. Ageing is also accompanied by all sorts of disorders ranging from simple to complex neurological degradation. In the past few decades, there has been a surge of knowledge of chemotherapeutic and curative properties of polyphenols, which are antioxidants present in plants and fruits to prevent the damage brought about by oxidative reactions. Though many reports have been published on herbal plants and meticulous intake of these bioactive compounds have been shown to protect against oxidative detriment lowering the risk of chronic and debilitating diseases (Hung et al., 2004), yet no innocuous permanent health and longevity supplement has been discovered.
Date palm (Phoenix dactylifera L. Arecaceae) constitutes a staple food for people of arid and semi-arid regions of the Middle East. The fleshy pericarp forms the pulp. A seed is usually oblong and forms about 10–15% of the fruit weight (Abdalla et al., 2012). With 7.9 million tons of date production in 2014, approximately 800,000 tons of date-pits were produced (FAO, http://www.fao.org). This huge amount of waste product could serve as a resource for the food industry and provide economic security to disadvantaged populations. Date seed have a higher percentage of protein and fat as compared to its pulp (Rahman, 2007). The protein fraction of the pits contains the preponderance of essential amino acids (Al-shahib and Marshall, 2003). Date seed is also rich in dietary fibers with 77.8–80.2 g/100 g fresh weight (Al-farsi and Lee, 2017). Its oil has a high UV absorbing potential thus making it a good option in the cosmetic world (Aris et al., 2014). To date no reports have been published regarding its role in the longevity of any organism. The presence of high flavonoid and phenolic content makes it a suitable candidate in the search for anti-aging products.
Syringic acid (SA), one of the major phenolics in date seed, synthesized via the shikimic acid pathway has demonstrated antihyperlipidemic, anticancer, nephroprotective, antimutagenic and antimicrobial activities in rat models (Cheemanapalli et al., 2018). These effects have also been shown with date consumption (Fa and Mm, 2013; Ahmed et al., 2016; Verma and Gautam, 2016; Khan et al., 2017). Supplement of 2–4% date palm to the diet of transgenic Alzheimer’s model mice has shown to decrease aggregation of the amyloid-beta protein and increase cognitive function (Selvaraju et al., 2015). The reported commendatory effects of date consumption may be attributed to the syringic acid present within it. Syringic acid is also abundantly present in pumpkin, olives, red wine, honey and acai palm.
Until the present there has been no report on the study of date palm pulp or its seed extract and syringic acid on C. elegans. Here, we used the date palm seed extract and syringic acid as its active ingredient on Caenorhabditis elegans, a nonparasitic, self-fertilizing hermaphrodite nematode. C. elegans is a good experimental model to study aging as it has a short lifespan and its genome can be easily manipulated under laboratory conditions. The process of aging in C. elegans is quite conserved and has similar deterioration in health with age like decreased cognitive abilities, decreased reproductive capacity, decreased motility and increased accrual of metabolic wastes as in humans (Iwasa et al., 2010). C. elegans is, therefore, a good model for testing therapeutic compounds. The effect of date seed and syringic acid on lifespan and health span parameters were tested using wild type (N2 Bristol), null mutant of daf-16 (mu86) and DAF-16::GFP (CF1139) strains.
2 Materials and methods
2.1 Chemicals and reagents
Syringic acid (SA) and sodium azide were purchased from HiMedia. Paraquat dichloride hydrate and (-)-epigallocatechin gallate (EGCG) were obtained from Sigma-aldrich. Reagents for extraction were of analytical grade and purchased from Merck.
2.2 Preparation of seed extract
Seeds of P. dactylifera (tamer stage of Medjool variety) were removed from the date palm fruits received as a gift, soaked in water and washed to detach any adhered date flesh. They were air dried at room temperature and ground into fine powder. One gram powder suspended in 40 ml of methanol (50% concentration) was used for extract preparation. The extract was prepared at room temperature for 72 h, with decanting the extract off every 24 h and resuspending the residue in fresh solvent. The filtrate collected was concentrated in a rotatory evaporator at 45 °C for 3 h and then dried through lyophilization. Different working concentrations of the P. dactylifera seed extract (PDS): 250 µg/ml, 500 µg/ml, 1000 µg/ml and 2000 µg/ml were made from a stock of 50 mg/ml of PDS and stored at 4 °C.
2.3 Reversed phase high performance liquid chromatography of the extract
HPLC conditions: Phenolic compounds of P. dactylifera seed extract were determined by reversed-phase high-performance liquid chromatography (RP-HPLC). Quantitative and qualitative validation of the phenolic compounds was done with a 1260 Quat pump, rheodyne manual injector, G1316A column compartment and 1260VWD detector by Agilent technologies with Agilent Openlab (ezchrome) software. Elution was done at 30 °C on a C-18 reversed phase column (250 × 4.6 mm, 5 µm particle size) of Agilent. The mobile phase was prepared by mixing methanol and water (of HPLC grade) in the ratio 1:4. 2 ml of acetic acid was added for better peak resolution. Isocratic elution was maintained at a flow rate of 1.0 ml/min. The apparatus was run at a wavelength of 280 nm.
Standard and sample preparation: Different phenolics for reference standards (>95% purity) were purchased from HiMedia. Different concentrations were prepared from their standards stock by dilution using the mobile phase as the diluents. The phenolics and the sample of crude extract of date seed powder was then filtered through a 0.45 µm Millipore filter before injecting into the pump. Injection volume of the phenolics was 20 µL.
Peaks were discerned by the similar retention time of the standards and that in the extract and by comparing the relative retention times as reported by Al Harthi et al (Al Harthi et al., 2015).
2.4 Total phenolic and flavanoid content (TPC and TFC) in P. dactylifera seed extract
The total phenols present in the methanolic extract of date seed was determined by Folin-Ciocalteu reagent method as previously described by Singleton et al. (1999). Gallic acid of concentration 100–500 mg/mL was used as standard to measure the total phenolic compounds. TPC was expressed as mg of gallic acid equivalents (GAE) per 100 g of date seed powder. The total flavonoids present in the methanolic extract of date seed were determined by aluminum chloride reagent method as previously described by Al-Farsi and Lee (2008). Catechin of concentration 100–500 mg/mL was used as standard to calibrate the measurements. TFC was expressed as mg of catechin equivalents (CAE) per 100 g of date seed powder. All the readings were taken three times and the mean value was used for calculation.
2.5 In vitro antioxidant assays
The antioxidant activities of methanol extract of date seed and syringic acid were evaluated using FRAP (ferric reducing antioxidant power) assay and hydrogen peroxide scavenging activity. The experiment was set up in triplicates and the optical density (OD) measurement was done through UV–VIS spectrophotometer.
2.5.1 Antioxidant activity using FRAP assay
The FRAP assay was carried out following the method developed by Benzie and Strain (1996). This assay measures the reducing power by measuring the reduction of ferric ion (Fe3+) to ferrous ion (Fe2+) in the presence of the antioxidant molecule. The OD of the intense blue color developed at low pH by the reduction of ferric tripyridyltriazine [Fe(III)-TPTZ] to ferrous tripyridyltriazine [Fe(II)-TPTZ] complex was measured at 593 nm wavelength. FeSO4 solution in the range 100 µM − 500 µM was used as a standard for calibration. The FRAP values of the date seed extract and syringic acid was expressed in terms of µmol Fe(II)/g dry mass.
2.5.2 Hydrogen peroxide scavenging activity
The H2O2 scavenging ability of the date seed extract and syringic acid was assessed following the method previously described by Ruch et al. (1989). The absorbance reading was taken at 230 nm wavelength. The scavenging of H2O2 was calculated using the formula: where
Ab = Absorbance of the blank (control)
As = Absorbance of the sample
2.6 Strains and maintenance
Different strains of Caenorhabditis elegans, the wild type N2 (Bristol), CF1038 (daf-16(mu 86)) and CF1139 (DAF-16::GFP) were procured from the Caenorhabditis Genetics Center (University of Minnesota, USA). All the strains were cultured and maintained on nutrient growth media (NGM) seeded with a spot of uracil auxotroph mutant of Escherichia coli OP50 at 20 °C in an incubator (Brenner, 1974).
2.7 Treatments
For lifespan assays and toxicity testing, age synchronized L1 stage worms were treated with different concentrations of PDS extract: 250, 500, 1000 and 2000 µg/mL (PDS); and syringic acid (SA): 200, 400, 600 and 800 µg/mL. For stress tolerance, pharyngeal pumping assay and florescence spectroscopy, 250 µg/mL PDS and 600 µg/mL SA treatment was given. For brood size assays, a dose of 250, 500 and 1000 µg/mL PDS and 200, 400 and 600 µg/mL SA was used as the treatment. The control group had 1% (v/v) methanol and the positive control group was obtained by treatment with 25 µg/mL EGCG.
2.8 Toxicity assessment and lifespan assay
For testing the toxic concentration and subsequent lifespan analyses of the N2 strain, 35 L1 worms were transferred onto live E. coli spotted NGM plates having the above mentioned concentrations of PDS and SA and counted as day 0. After the worms reached L4 stage, they were picked and transferred daily till reproduction ceased and then transferred every second day to fresh treated Petri plates to avoid progeny count. The live worms were transferred onto fresh Petri plates and the worms which failed to show any response in response to prodding were scored dead. The counting was carried on till all the worms were dead or censored. A similar procedure was followed for the daf-16 (mu86) strain with non-toxic concentration of the sample which showed maximum lifespan in N2. Three replicates for each concentration were analyzed. Kaplan-Meier survival curves were made and data analysis was done through Log-rank (Mantel-Cox) test using Graphpad Prism 5.
2.9 Survival assay under paraquat and H2O2 induced oxidative stress
2.9.1 Paraquat stress assay
Paraquat is a generator of superoxide radicals which have a damaging effect on the lipid membrane. A viability experiment in the presence of oxidative stress due to paraquat was performed following the method previously described by Ishii et al (Ishii et al., 1990). 50 day 2 adult N2 worms were transferred to the culture media containing 10 mM paraquat with a platinum wire. The plates were incubated at 20 °C for 10 h followed by examining the survivors and scoring the dead.
2.9.2 H2O2 stress assay
H2O2 is a weak oxidizing agent and brings about damage by generating hydroxyl radicals. Day 2 adult N2 worms were washed 3–4 times with chilled M9 buffer followed by soaking them in 10 mM hydrogen peroxide in PBS for 4 h. 50 worms per plate were transferred to the culture media and incubated overnight at 20 °C. Next day the counting was done and the worms which showed no movement were scored as dead.
2.10 Survival assay under thermal stress
Adult N2 worms with about 50 pretreated worms per plate were incubated at 32 °C for 4 h. Counting of the surviving worms was done on an hourly basis. The worms which did not move even after prodding slightly with a platinum wire were scored as dead.
2.11 Pharyngeal pumping assay
With age there is a progressive decline in muscular functioning manifested by a decrease in pharyngeal pumping rate. This leads to a decrease in food intake mimicking a calorie restriction like effect. To determine this effect, the pharynx movement of pretreated N2 worms was counted at a time lapse of 15 s over a period of 1 min under a Radical® stereomicroscope. The counting was done on days 5, 8, 10 and 12. Three independent sets of experiments were performed.
2.12 Brood size assay
Synchronized L4 worms of N2 were used to measure the brood size. Three plates having 25 worms each were set up for each concentration. After the egg laying started, worms were transferred to fresh and treated plates every 24 h until egg laying ceased. The total numbers of larva hatched out from the eggs of each worm on each plate were counted.
2.13 Fluorescence microscopy
DAF-16::GFP nuclear localization assay
A DAF 16 nuclear localization assay was performed on age synchronized L4 staged DAF-16::GFP transgenic C. elegans. About 20 L4 worms from each treatment were picked and mounted on a 2% agarose pad and covered with a glass slide. The worms on the slide were imaged by florescence microscopy under 10× magnification. For examination of the GFP transgenic lines, the filters used for excitation and emission were 400/30 and 508/20 nm, respectively.
2.14 Statistical analyses
All the experiments were done in triplicate. Graphs were made using Prism 5 (GraphPad Software, San Diego, CA). Comparisons between two groups were done by paired Student’s t-test while that between multiple groups were done by two- way analysis of variance (ANOVA) followed by analysis with a Bonferroni test.
3 Results
3.1 Total phenolic and flavanoid contents of date seed extract
The total phenolic and flavanoid contents in the crude extract of P. dactylifera seed was found to be 449 mg gallic acid equivalent (GAE) and 458 mg catechin equivalent (CE) per 100 g of date seed powder, respectively. HPLC-VWD chromatograms of the date seed extract at 280 nm are shown in Fig. 1. Quantitative data are listed in Table 1. Gallic acid was the main compound in the extract (19.53 µg/mL). Other phenolics were syringic acid (10.72 µg/mL), p-coumaric acid (3.598 µg/mL) and caffeic acid (4.007 µg/mL). The retention time of the phenolics lied between 4 min to 20 min and was in the following sequence: Gallic acid < p-Coumaric acid < Caffeic acid < Syringic acid.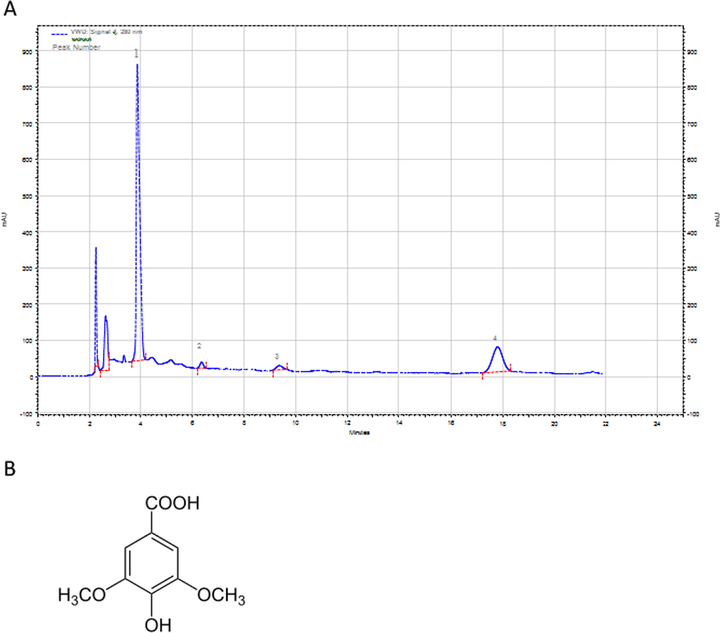
(A) HPLC-VWD chromatogram of date seed extract at 280 nm. Peaks are of compound gallic acid (1), p-coumaric acid (2), caffeic acid (3) and syringic acid (4). (B) Structure of syringic acid.
Phenolic
Retention time (minutes)
Concentration (µg/mL)
Gallic acid
3.86
19.53
p-Coumaric acid
6.348
3.598
Caffeic acid
9.7
4.007
Syringic acid
17.5
10.72
3.2 In vitro antioxidant activities of date seed extract and syringic acid
The crude extract and syringic acid demonstrated high antioxidant activities in a concentration dependent manner. The reducing power (FRAP) of date seed extract showed low activity of 41.26 ± 1.154 µmol Fe (II)/g dry mass at 250 µg/mL while high activity of 199.60 ± 1.134 µmol Fe (II)/g dry mass at 2000 µg/mL. Similar trend was observed with syringic acid. There was an increase of 129.28% in the reducing capability with low to high concentration of syringic acid (Fig. 2). Regarding the H2O2 scavenging ability, the present study showed that the date seed and syringic acid capable of removing H2O2 from the reaction mixture are also in concentration dependent fashion. The highest level of PDS (2000 µg/mL) showed the greatest potential to nullify the action of H2O2. Syringic acid also showed maximum antioxidant potential at highest test level (800 µg/mL) (Table 2). Also, the antioxidant activity of the crude extract showed linear co-relation with the total phenolic content (R2 = 0.9965 for FRAP and 0.9876 for H2O2 scavenging activity). Each value is expressed as mean ± SEM.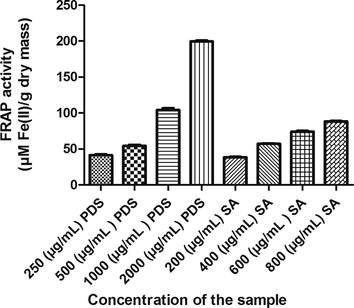
Antioxidant (FRAP) activity of P. dactylifera seed extract and Syringic acid.
Sample (µg/mL)
FRAP activity (µmol Fe(II)/g dry mass)
H2O2 scavenging activity (%)
Date seed extract
250
41.26 ± 1.154
27.51 ± 0.849
500
54.36 ± 1.455
37.05 ± 0.494
1000
104.10 ± 2.413
45.06 ± 0.829
2000
199.60 ± 1.134
76.21 ± 0.592
Syringic acid
200
38.46 ± 0.575
6.70 ± 0.157
400
57.26 ± 0.614
10.43 ± 0.326
600
73.96 ± 1.193
33.98 ± 0.172
800
88.18 ± 0.594
51.43 ± 0.847
3.3 Date seed and syringic acid increases longevity in C. elegans
The age-synchronized N2 strain of C. elegans was fed with different concentrations of PDS extract and syringic acid for the whole lifespan. The results show that PDS extract treatment increases lifespan at lower concentrations. Doses beyond 1000 µg/mL exhibited toxicity. The increment in mean lifespan of N2 at 250 µg/mL PDS extract was 18.50% (p < 0.001). Doses above 250 µg/mL comparatively decrease lifespan (Fig. 3A). There was significant 11.90% (p < 0.05) decline in lifespan at 2000 µg/mL PDS. Syringic acid increased the lifespan in a dose dependent manner with maximum lifespan being demonstrated at 600 µg/mL (Fig. 3B). The mean lifespan was significantly extended by 14.40% (p < 0.001) with 600 µg/mL SA (Table 3). At a concentration above 600 µg/mL, the lifespan decreased significantly by 15.24% (p < 0.005). Thus, the isolated secondary metabolite syringic acid demonstrated lethality about three-fold as compared to the crude extract of date seed. A decrease in pharyngeal pumping can have dietary restriction-like effects thus increasing lifespan (Hekimi and Lakowski, 1998). Consequently, we hypothesized that PDS treatment could lead to a decrease in pharyngeal pumping rates. To test this hypothesis, we noted the pharyngeal pumping rates and found that they significantly increased by 92.84% and 59.30% with 250 µg/mL PDS and 600 µg/mL SA treatment, respectively, as compared to control (p < 0.001) (Fig. 3C). This indicated that the increase in longevity is not due to dietary restriction, rather the treatment increased the muscle function.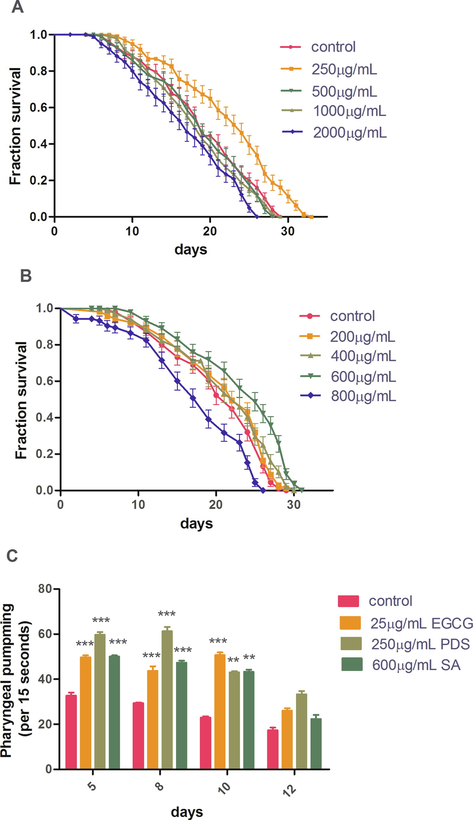
PDS treatment of 250 µg/mL concentration (A) and SA treatment of 600 µg/mL (B) extend the life span of N2 (p < 0.001) with a significant increase in the pharyngeal pumping at that concentration (C).
Genotype
Treatment
Mean ± SEM
N
p-value
N2 (wild type)
Control
18.49 ± 0.6656
90
–
N2 (wild type)
250µg/mL PDS
21.91 ± 0.7087
90
0.0006
N2 (wild type)
500µg/mL PDS
18.27 ± 0.6490
92
0.8109 (NS)
N2 (wild type)
1000µg/mL PDS
18.09 ± 0.6199
91
0.6654 (NS)
N2 (wild type)
2000µg/mL PDS
16.29 ± 0.6394
92
0.0181
N2 (wild type)
Control
20.14 ± 0.6272
95
–
N2 (wild type)
200µg/mL SA
20.59 ± 0.6464
96
0.6126 (NS)
N2 (wild type)
400µg/mL SA
20.89 ± 0.6601
94
0.4069 (NS)
N2 (wild type)
600µg/mL SA
23.04 ± 0.6431
94
0.0014
N2 (wild type)
800µg/mL SA
17.07 ± 0.6869
96
0.0012
daf 16 (mu86)
Control
17.48 ± 0.6166
94
–
daf 16 (mu 86)
250µg/mL PDS
19.70 ± 0.6141
91
0.0114
daf 16 (mu 86)
600µg/mL SA
19.34 ± 0.7097
93
0.0485
3.4 Date seed and syringic acid increases stress tolerance in C. elegans
Age-synchronized L1 larvae of the N2 strain treated with PDS extract and SA were subjected to oxidative stress generated by paraquat and H2O2 and thermal stress of 32 °C. The results show that PDS extract and SA pretreated worms have augmented survival rate in the stress condition as compared to the untreated worms. The survival of the worms significantly increased by 69.07% (p < 0.001) with 250 µg/mL PDS extract when compared to the control (1% methanol + 10 mM paraquat) under paraquat stress. The corresponding effect of SA was also significantly increased by 58.17% (p < 0.001) at 600 µg/mL SA. The survival rate was similar to the positive control EGCG (60%) (Fig. 4A). Significant increment (p < 0.05) in survival rate was observed under H2O2 stress by 55.4% and 48.65% with 250 µg/mL PDS extract and 600 µg/mL SA, respectively, when compared to the control (1% methanol + 10 mM H2O2). This result showed similarity to the positive control EGCG (78.67%) (Fig. 4B). Upon thermal stress of 32 °C, the survival of worms fed on 250 µg/mL of PDS extract increased by 71.6% (n = 142, p < 0.05). SA also significantly (p < 0.01) provided resistance against thermal stress by 37.26% (n = 145) at 600 µg/mL when compared to the control. The resistance provided by EGCG showed similar result (47.86%) (Fig. 4C).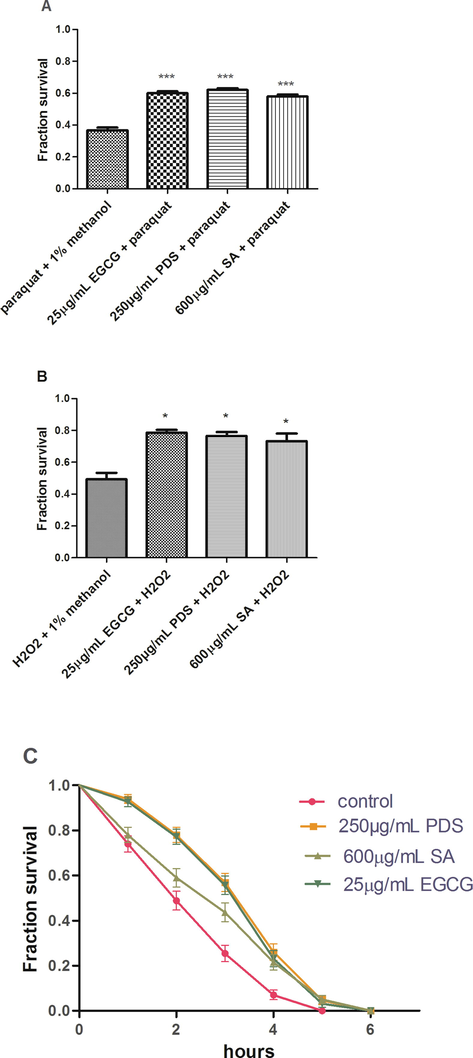
SA and PDS protect the N2 strain of C. elegans against oxidative and thermal stress. (A) Fraction survival of worms against oxidative stress induced by 10 mM paraquat. The mean number of worms surviving the paraquat stress at 250 µg/mL PDS and 600 µg/mL SA were 31.00 ± 0.577 and 29.00 ± 0.577, respectively, compared to the control (18.33 ± 0.882) and positive control 25 µg/mL EGCG (30.00 ± 0.577). (B) The mean number of worms that surviving significantly (p < 0.05) under 10 mM H2O2 stress was 38.33 ± 1.202 at 250 µg/mL PDS and 37.67 ± 2.404 at 600 µg/mL SA compared to the control (24.67 ± 2.028) and positive control 25 µg/mL EGCG (39.33 ± 0.882). (C) A Kaplan Meier survival curve was made and analyzed through Log-rank (Mantel-Cox) test when worms were subjected to thermal stress of 32 °C. The mean numbers of worms surviving significantly were 24.60 ± 8.33 at 250 µg/mL PDS and 19.67 ± 6.286 at 600 µg/mL SA, respectively, vs. control 14.33 ± 6.225 and positive control 23.93 ± 8.291. Data are expressed as Mean ± SEM. *p < 0.05, ***p < 0.001.
3.5 Date seed and syringic acid profoundly increases brood size
The increase in fecundity was counted by observing the number of larva hatched after treatment. The fertility was increased by 117.9% and 50% at 500 µg/mL, and 1000 µg/mL PDS extract, respectively (p < 0.01) and by 100.6% and 46% at 200 µg/mL and 400 µg/mL SA, respectively (p < 0.01) (Fig. 5).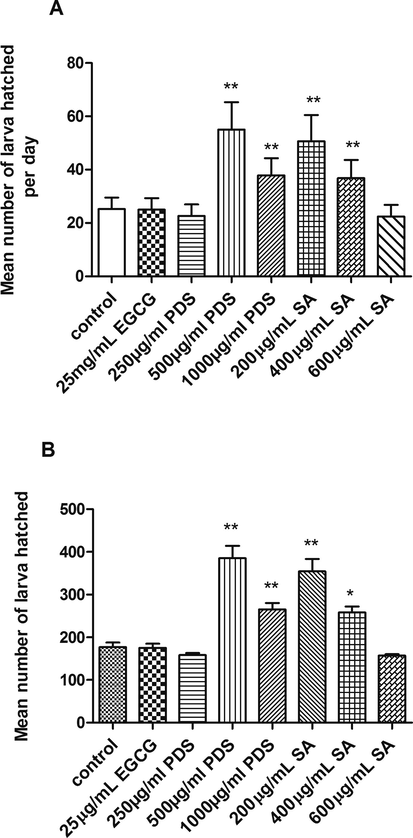
The brood size of the worms increased upon PDS and SA treatment (**p < 0.01).
3.6 Date seed extract and syringic acid induced alteration in lifespan and stress tolerance in C. elegans is independent of the DAF-16 nuclear localization
The insulin signaling pathway is highly conserved in humans and worms. DAF-16 (the fork headed like transcription factor homologue of the mammalian FoxO protein) is the central protein which regulates gene expression responsible for several biological processes through nuclear localization. To determine the action of PDS and SA on the Insulin/Insulin like Growth Factor-1 signaling (IIS-1) pathway, we used strains CF1139 (DAF-16::GFP) and CF1038 (daf-16 null mutant mu86). DAF-16::GFP is a transgenic animal in which nuclear localization of DAF-16 can be visualized by subjecting the worms to heat stress for 15 min. PDS and SA treatment to DAF-16::GFP showed no changes in the florescence pattern when compared to the control DAF-16::GFP worms unlike EGCG treatment which caused nuclear localization (Fig. 6C). This result signified that both exert their effects independent of the DAF-16 translocation. To further explore this result, we examined the lifespan increment and thermotolerance in long lived daf-16 null mutant mu86. We exposed the CF1038 strain with 250 µg/mL PDS extract and 600 µg/mL SA and found no significant difference in the mean lifespan and also in the mean number of worms surviving thermal stress when compared to the N2 strain. The lifespan increased by 12.70% on 250 µg/mL PDS and 10.64% on 600 µg/mL SA treatment (control mean lifespan = 17.48 ± 0.617 vs. 19.70 ± 0.614 for PDS and 19.34 ± 0.710 for SA, p < 0.05) (Fig. 6A). The mean survival under thermal stress increased by 73.84% with PDS and 46.11% with SA (PDS mean ± SEM = 22.13 ± 8.261, SA mean ± SEM = 18.60 ± 7.425 vs. control mean ± SEM = 12.73 ± 6.004, p < 0.0001). The survival of EGCG treated worms showed no significant difference from the control, mean ± SEM = 13.27 ± 6.017 (Fig. 6B).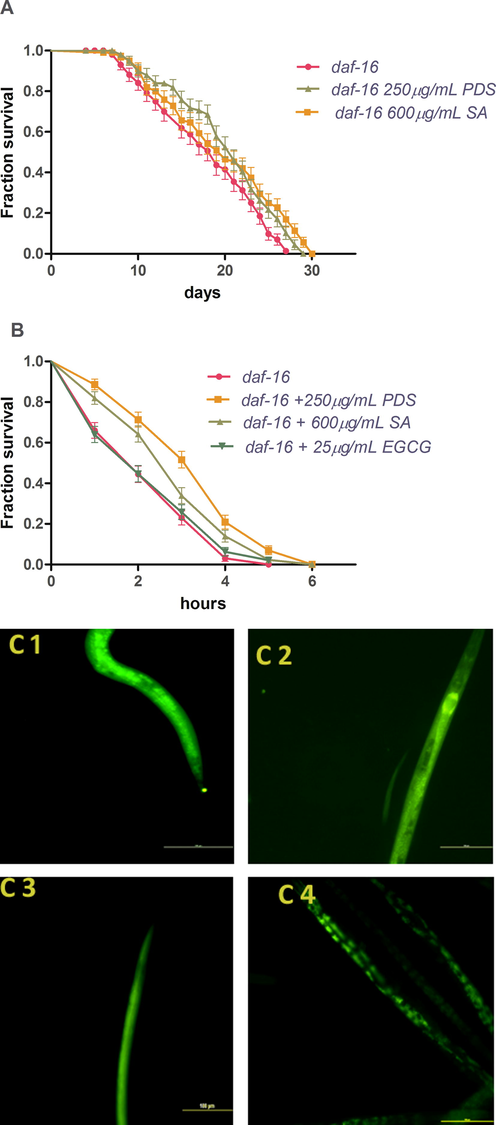
Survival curve of the effect of PDS and SA treatments on the lifespan (A) and thermal stress (B) of daf-16 mutant worms. Fluorescent images of DAF-16::GFP showing cytoplasmic localization of DAF-16 in control (1% methanol) (C1), PDS (C2) and SA (C3) treatment and nuclear localization with EGCG treatment(C4). Images show scale bar of 100 µm.
4 Discussion
Ageing can be considered as a cumulative decline in cellular activities over the lifetime. It is associated with progressive regression of homeostasis and physiological functions of the cells and thus affecting the whole organism. Studies in various model organisms support the hypothesis that generation of ROS and concomitant oxidative damage increases with age (Stadtman, 2006). The polyphenol and flavonoid molecules present in various plant sources act as antioxidants to curb the damage brought about by ROS to a certain extent. We checked the efficacy of ployphenols present in date seed as a potential candidate for delaying the effects of ageing. The content of the phenolics and flavonoids may vary depending upon the stage of ripening and harvesting of the fruit. They also depend on the climatic condition of the growth of the plant. The physicochemical analysis of the extract by reverse phase high performance liquid chromatography (RP-HPLC) demonstrated the presence of 4 major phenolic compounds which might be acting as antioxidants. The total phenolic and flavanoid contents in the seed extract was much higher as that reported by El-Mergawi et al. (2016). The isolated secondary metabolite also showed in vivo antioxidant activities along with the crude extract. Several reports have previously been published about the beneficial effect of date seed on a rat model and Drosophila (Takaeidi et al., 2014).
The organism used in our study is Caenorhabditis elegans chosen due to its various advantages over other model organisms. Different signaling pathways like TOR, Insulin/IGF-1 and germline signaling pathways are highly conserved among mammals and C. elegans (Lapierre and Hansen, 2012). Moreover, there is also 83% homology in the proteome of C. elegans and human genes (Lai et al., 2000) permitting the researchers to intrigue the molecular basis of ageing. Our results revealed that exposure to date seed extract preferentially promotes healthy ageing.
Various physiological functions like pharyngeal pumping, body bending, sarcopenia and lipofuscin accumulation are considered as hallmarks of ageing in C. elegans. Contrary to the established fact that reduction in pharyngeal pumping increases lifespan, our treatment increased the lifespan along with increase in the pharyngeal pumping rate showing that the increase in lifespan is not due to caloric restriction. The treatment also increased the fecundity and enhanced stress tolerance· H2O2 and paraquat are superoxide radical generators. When worms are subjected to a high dose of these stressors, the resulting intracellular toxicity by ROS is prevented by syringic acid as shown by the increased survival rate. Surprisingly, these effects were more pronounced in the treatment with crude seed as compared to the purified component. Earlier reports have shown that the effect of active compound isoliquitigenin from Glycyrrhiza uralensis has more potent effect on ROS quenching in C. elegans than the plant extract (Link and Wink, 2019). The present result may be due to the additive or synergistic effects of different phenolics and flavonoids present in the seed. Gallic acid which is the most abundant phenolic in the date seed has previously been shown to increase the lifespan in C. elegans (Saul et al., 2011). The pathways inducing stress-response genes and affecting lifespan in C. elegans have been identified and these also show homology with human genome. DAF-16 and SKN-1, homologues of vertebrate FOXO and Nrf proteins of IIS-1 and nuclear factor erythroid 2-related factor 2 pathway, respectively, promotes expression of lifespan extending and stress tolerance genes. The increment in lifespan and stress tolerance upon date seed and syringic acid treatment might be through action on any of these genes, so we examined whether the effect of mutation in one of the gene (daf-16) impaired the ability to show similar response as in wild type N2 or not. The premise of hypothesis of the experiment was that our treatment would significantly increase lifespan and promote stress tolerance in daf-16 mutant worms as compared to N2 as was observed in previous studies of Duangjan et al. (2019). Treating the daf-16 mutant worms with date seed extract and syringic acid, lifespan increased by 12.70% and 10.64%, respectively, but this increase was not statistically significantly different as compared to that in N2 strain. Also, the effect of thermal stress in daf-16 null mutants was not different to that in N2 strains. These observations indicated the action to be independent of DAF-16. To further discern the non-involvement of DAF-16 we used green florescence tagged DAF-16 (DAF-16::GFP) transgenic strain. There was no cytoplasmic to nuclear translocation of DAF-16 upon PDS and SA treatment unlike previous established results of treatment with ECGC. One interpretation of our findings is that PDS and SA were exerting its effect independent of DAF-16 translocation. The effect may be due to some other genes via different pathway involved in lifespan extension and induction of stress tolerance. Or it may be acting on some other protein downstream in IIS-1 pathway. Lifespan extension and increase in stress tolerance have been previously shown by crude extract of blueberries to be independent of both DAF-16 and SKN-1 (Wilson et al., 2006). On contrary, leaf extract of Anacardium occidentale mediated resistance to oxidative stress through both DAF-16 and SKN-1 (Duangjan et al., 2019). Thence, further investigation for PDS and SA effect through SKN-1/Nrf-2 pathway needs to be investigated. Date has long been used in contemporary medicine for the treatment of infertility. The phytochemical compounds like genistein, selenium and vitamin A present in the fruit are known to protect sperm membrane and maintain sperm count. Phytosterols and phytoestrogens also regulates estrogen imbalance in the body (Saryono et al., 2018). The generation of free radicals in the body also affects oogenesis and embryonic development. The polyphenols in the date fruit and its seed quenches the free radicals and boost fertility. Worms fed on a high concentration of date seed extract (>250 µg/mL) have shown to have increased egg laying capacity. In our study, syringic acid also demonstrated an increase in brood size, suggesting that it might also be responsible for this effect in the crude treatment to some extent. This observation suggests the action of date seed might be both on the sperm and oocyte development.
In conclusion, syringic acid had demonstrated high antioxidative property in other animals and a similar effect has now been seen in worms as well. Therefore, regular consumption of date along with its seed in powder form can boost health and can now be considered as a nutritional supplement.
Acknowledgements
Special thanks to the Caenorhabditis Genetics Center for supplying the worm strains for this work. S Bhushan and M Haque kindly helped in data analysis. A.J. thanks the UGC- Maulana Azad National Fellowship for financial support.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Physico-chemical characteristics of date seed oil grown in Sudan. Am. J. Appl. Sci.. 2012;9:993-999.
- [CrossRef] [Google Scholar]
- Anti hyperlipidemic and hepatoprotective effects of native date fruit variety “Aseel” (Phoenix dactylifera) Pak J Pharm Sci.. 2016;26:1945-1950.
- [Google Scholar]
- Quantification of phenolic compounds, evaluation of physicochemical properties and antioxidant activity of four date (Phoenix dactylifera L.) varieties of Oman. J. Taibah Univ. Med. Sci.. 2015;10:346-352.
- [CrossRef] [Google Scholar]
- Optimization of phenolics and dietary fibre extraction from date seeds. Food Chem.. 2008;108(3):977-985.
- [CrossRef] [Google Scholar]
- Nutritional and functional properties of dates: a review. Crit. Rev. Food Sci. Nutr.. 2017;48:877-887.
- [CrossRef] [Google Scholar]
- The fruit of the date palm: its possible use as the best food for the future ? Int. J. Food. Sci. Nutr.. 2003;54:247-259.
- [CrossRef] [Google Scholar]
- Solubility of phoenix dactylifera seed oil in supercritical carbon dioxide (Sc-Co 2) using empirical model. Key Eng. Mater.. 2014;594–594:301-305.
- [CrossRef] [Google Scholar]
- The ferric reducing ability of plasma (FRAP) as a measure of ‘Antioxidant Power': The FRAP Assay. Analytical Biochem.. 1996;239:70-76.
- [Google Scholar]
- Biomedicine & Pharmacotherapy Syringic acid (SA) - A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother.. 2018;108:547-557.
- [CrossRef] [Google Scholar]
- Lifespan extending and oxidative stress resistance properties of a leaf extracts from Anacardium occidentale L. in Caenorhabditis elegans. Oxidative Med. Cellular Longevity.. 2019;19
- [CrossRef] [Google Scholar]
- Phenolic profiles and antioxidant activity in seeds of ten date cultivars from Saudi Arabia. J. Food, Agric. Environ.. 2016;14(2):38-43.
- [Google Scholar]
- Antibacterial activity of date palm (PhoenixDectylifera L.) fruit at different ripening stages. J. Food Process Technol.. 2013;4
- [CrossRef] [Google Scholar]
- The genetics of caloric restriction in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U S A. 1998;95:13091-13096.
- [CrossRef] [Google Scholar]
- Fruit and Vegetable Intake and Risk of Major Chronic Disease. J. Natl. Cancer Inst.. 2004;96:1577-1584.
- [CrossRef] [Google Scholar]
- A methyl viologen-sensitive mutant of the nematode Caenorhabditis elegans. Mutat. Res.. 1990;237:165-171.
- [CrossRef] [Google Scholar]
- Novel EGF Pathway Regulators Modulate C. elegans Healthspan and Lifespan via EGF Receptor, PLC-γ and IP3R Activation. Aging Cell. 2010;9:490-505.
- [CrossRef] [Google Scholar]
- Anti-cancer effects of Ajwa dates (Phoenix dactylifera L.) in diethylnitrosamine induced hepatocellular carcinoma in Wistar rats. BMC Complement Altern Med.. 2017;17:418.
- [CrossRef] [Google Scholar]
- Identification of Novel Human Genes Evolutionarily Conserved in Caenorhabditis elegans by Comparative Proteomics. Genome Res.. 2000;10:703-713.
- [CrossRef] [Google Scholar]
- Lessons from C. elegans: signaling pathways for longevity. Trends Endocrinol. Metab.. 2012;23:591-598.
- [CrossRef] [Google Scholar]
- Isoliquirtigenin exerts antioxidant activity in Caenorhabditis elegans via insulin-like signaling pathway and SKN-1. Phytomedicine. 2019;55:119-124.
- [CrossRef] [Google Scholar]
- Composition characterisation and thermal transition of date pits powders. J. Food Eng.. 2007;80:1-10.
- [CrossRef] [Google Scholar]
- Prevention of cytotoxicity and inhibition of intracellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogenesis. 1989;10:1003-1008.
- [CrossRef] [Google Scholar]
- Effects of dates fruit (Phoenix dactylifera L.) In the female reproductive process. Int. J. Recent Adv. Multidisciplinary Res.. 2018;3:1630-1633.
- [Google Scholar]
- Diversity of polyphenol action in Caenorhabditis elegans: between toxicity and longevity. J. Nat. Prod.. 2011;74:1713-1720.
- [CrossRef] [Google Scholar]
- Diet rich in date palm fruits improves memory, learning and reduces beta amyloid in transgenic mouse model of Alzheimer ’ s disease. J. Ayurveda Integr. Med.. 2015;6:111-120.
- [CrossRef] [Google Scholar]
- [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagents. Methods Enzymol.. 1999;299:152-178.
- [CrossRef] [Google Scholar]
- Protein oxidation and aging. Free Radical Res. Free Radic. Res.. 2006;40:1250-1258.
- [CrossRef] [Google Scholar]
- The effect of date seed (Phoenix dactylifera) extract on paraoxonase and arylesterase activities in hypercholesterolemic rats. Jundishapur J. Nat. Pharm. Prod.. 2014;9(1):30-34.
- [Google Scholar]
- Antimutagenic potential of date palm (Phoenix dactylifera) fruit aqueous extract in suppressing induced mutagenesis and purification of its bioactive constituent. MOJ Food Process Technol.. 2016;2:179-185.
- [CrossRef] [Google Scholar]
- Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell. 2006;5:59-68.
- [CrossRef] [Google Scholar]




