

Translate this page into:
The extraction, antioxidant and against β-amyloid induced toxicity of polyphenols from Alsophila spinulosa leaves
⁎Corresponding author. yuanming@sicau.edu.cn (Ming Yuan)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Alsophila spinulosa is a tree-like fern, and many evidences suggested that plant polyphenols had the potential therapeutic for Alzheimer s disease (AD). Herein, polyphenols (ASP) was isolated from A. spinulosa leaves and its major constituent were isoorientin and vitexin. ASP displayed excellent antioxidant activity and obvious anti-lipid peroxidation capacity in vitro. ASP improved the survival rate of C. elegans under high temperature by enhancing the antioxidant enzymes activities and decreasing the lipid peroxidation level. Moreover, ASP alleviated β-amyloid (Aβ) induced paralysis and reduced Aβ deposition, decreased reactive oxygen species (ROS) accumulation and improved the level of skn-1 mRNA. In addition, ASP decreased the levels of pdk-1 and akt-1 mRNA in P13K/AKT signaling pathway. In conclusion, ASP may be a potential ingredient for the alleviation of AD.
Keywords
Alsophila spinulosa leaves
Polyphenols
Antioxidant
Alzheimer‘s disease
P13K/AKT

- AD
-
Alzheimer`s disease
- ASP
-
A. spinulosa leaves polyphenols
- Aβ
-
β-amyloid
- APP
-
Amyloid precursor protein
- ROS
-
Reactive oxygen species
- UAE
-
Ultrasonic assisted extraction
- RSM
-
Response surface methodology
- BBD
-
Box-Behnken Design
- DPPH
-
1,1-diphenyl-2-picrylhydrazyl
- ABTS
-
2,2-azino-bis(3- ethylbenzothiazoline-6-sulfonic acid)
- SOD
-
Superoxide dismutase
- CAT
-
Catalase
- MDA
-
Malondialdehyde
- NGM
-
Nematode growth medium
- HPLC
-
High performance liquid chromatography
- DCFH-DA
-
2`,7`- Dichlorodihydrofluorescein diacetate
- GSH
-
Glutathione
Abbreviations
1 Introduction
AD, representing 60%-80% of all dementia, occurred mainly over the age of 65, and it was a neurodegenerative disease related to memory and cognitive impairment (Grontvedt et al., 2018). Although its exact pathogenesis is uncertain, the hypothesis about Aβ cascade is considered the major pathological cause (Yang et al., 2019). The aggregation of Aβ will trigger neurotoxicity and oxidation (Shankar et al., 2008). One of the major features for AD is oxidative damage. Aβ entered mitochondria, which would promote the accumulation of ROS and induced the damage to the integrity of the cell structure in early course of AD (Pratico, 2008). Antioxidants might play a crucial part in prevention or mitigation of the symptom of AD (Petrovic et al., 2020). Some studies indicated that natural polyphenols had great potential in treatment of AD (Zeng et al., 2015; Freyssin et al., 2018).
Alsophila spinulosa (Hook.) Tryon (Cyatheaceae) is a tree-like fern (Morton, 1971). The stem of A. spinulosa is known as “Caulis Alsophilae” in China, and its a Chinese traditional medicine for the cough, asthma, cold and rheumatism. But its leaf is neglected and discarded. Polyphenols is a secondary metabolic product widely existing in plants and received considerable attention due to their health-promoting properties in neurodegeneration diseases, including AD and Parkinson’s disease (PD) (Abbas et al., 2016). Unfortunately, there are many polyphenols, such as vitexin, isoorientin, apigenin and luteolin in A. spinulosa leaves (Chen et al., 2008), but its mitigation effect of AD has not been reported.
The ultrasonic-assisted extraction (UAE) is considered as a green method for polyphenols extraction from the natural material because it could save energy, time, and solvents (Ameer et al., 2017). Therefore, UAE may be an efficient and energy-saving way to extract polyphenols from the leaves of A. spinulosa. Response surface methodology (RMS) is a scientific optimization method, and the Box–Behnken design (BBD) is a good design for RSM. They are widely used to optimize the extraction of polyphenols. Unfortunately, there was no report that extraction of polyphenols from A. spinulosa leaves by UAE and optimization with RSM.
In this work, the extraction polyphenols from A. spinulosa leaves was optimized with RSM and purified by AB-8 macroporous resin. The major constituent was analyzed by HPLC. In addition, their antioxidant activities and their potential effects on AD were investigated.
2 Materials and methods
2.1 1 Materials and chemicals
Sample leaf of A. spinulosa was gifted by Hongya Qingyuan Sci-tech Co., LTD (Hongya county, Sichuan Province, China) and identified by professor Ming Yuan. 1,1-diphenyl-2-picrylhydrazyl (DPPH), 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), and Folin-Ciocalteu were purchased from Solarbio Sci-tech Co., Ltd. (Beijing, China). The test kits of superoxide dismutase (SOD), catalase (CAT) and malondialdehyde (MDA) were obtained from Jiancheng Bioengineering Institute (Nanjing, China). The kits of RNAisoPlus reagent, RNA reverse transcription and fluorescent qPCR were purchased from Takara Biotechnology Co., Ltd. (Dalian, China). The phenolic standards (HPLC ≥ 98%) were obtained from Plant Origin Biological Co., Ltd. (Nanjing, China). Other reagents were purchased from Chengdu Kelong Chemical Industry. All reagents are analytically pure.
The C. elegans strains used in this study are N2 (wild type) and CL4176 (dvIs27 [myo-3p::A-Beta (1–42)::let-851 3′UTR) + rol-6(su1006)] X). The C. elegans CL4176, expressing human Aβ peptide in muscle cell, was gifted by Prof. Yunjiao Chen (South China Agricultural University, China). C. elegans was cultured on nematode growth medium (NGM) and seeded with Escherichia coli OP50 at 20 °C (N2) or 16 °C (CL4176). For experiments, the synchronous populations of CL4176 worms had cultured to the L1 stage and transferred to with or without ASP plates and incubated at 16 °C for 36 h, then moved to 25 °C for promoting the expression of Aβ.
2.2 RSM for the extraction of polyphenols
On the basis of single factors experiments, BBD with four key parameters (ultrasonic power, liquid–solid ratio, ethanol concentration and extraction time) at three levels was employed to optimize extraction parameters. Table 1 showed the 29 experiments which were designed by Design-Expert 8. The data of BBD were explained by the following second-order polynomial equation.
Where Y is the yield of polyphenols, Xi and Xj represents independent variable, and β0, βi, βii and βij represents regression coefficient. A1 means ultrasonic power, A2 means liquid–solid ratio, A3 means ethanol concentration, and A4 means extraction time.
Number
A1 (W)
A2 (mL/g)
A3 (%)
A4 (min)
Yield (%)
1
−1 (210)
−1 (20)
0 (40)
0 (20)
6.10
2
1 (270)
−1
0
0
5.86
3
−1
1 (30)
0
0
5.39
4
1
1
0
0
5.88
5
0 (240)
0 (25)
−1 (20)
−1 (15)
6.41
6
0
0
1 (60)
−1
6.88
7
0
0
−1
1 (25)
5.68
8
0
0
1
1
4.91
9
−1
0
0
−1
5.51
10
1
0
0
−1
8.19
11
−1
0
0
1
6.68
12
1
0
0
1
4.30
13
0
−1
−1
0
5.79
14
0
1
−1
0
5.55
15
0
−1
1
0
5.76
16
0
1
1
0
5.28
17
−1
0
−1
0
6.95
18
1
0
−1
0
6.35
19
−1
0
1
0
6.05
20
1
0
1
0
6.95
21
0
−1
0
−1
6.07
22
0
1
0
−1
5.67
23
0
−1
0
1
4.37
24
0
1
0
1
4.35
25
0
0
0
0
8.79
26
0
0
0
0
8.75
27
0
0
0
0
8.90
28
0
0
0
0
8.89
29
0
0
0
0
8.86
2.3 Extraction and purification of polyphenols
The leaves of A. spinulosa was washed and dried at 50 °C, then crushed and sifted through 100 mesh sieves and pre-treated with petroleum ether to remove pigments and lipids. The samples were extracted with 40% (v/v) ethanol (1:25, w/v) through an ultrasonic-assisted technique (240 W) for 20 min (Savic and Gajic., 2020). The supernatant was obtained by centrifugation 4000 rpm for 10 min, and then dried by freeze dryer.
The crude polyphenols was dissolved with ethanol, and purified by a macroporous resin column of AB-8 (Xi et al., 2015). 70% ethanol worked as eluent. The elution curve was drawn by the Folin-Ciocalteu method and the major peak was collected. Purified polyphenols were still named ASP and lyophilized.
2.4 Determination of total polyphenols
The total polyphenols was quantified according to Canalis et al. (2020). 0.05 mL ASP was added to 0.05 mL Folin-Ciocalteu and 0.15 mL 10% sodium carbonate and reacted in dark for 2 h. The absorbance of polyphenols was at 760 nm and the total polyphenols content was expressed as milligrams gallic acid equivalents per gram (mg GAE/g) according to the standard curve of gallic acid.
2.5 HPLC analysis
The component of ASP was analyzed using HPLC system (Agilent 1260, Agilent, Santa Clara, CA, USA) with an Agilent C18 reverse-phase column (5 μm, 250 mm × 4.6 mm) and an ultraviolet detector at 320 nm. 0.2% phosphoric acid solution (A) and acetonitrile (B) worked as mobile phase, and the gradient elution was: 0–25 min, 14%-40% B, 25–35 min, 40%-16% B, with 0.5 mL/min of the flow rate.
2.6 Assays of antioxidant activity in vitro
The scavenging activities of DPPH radical, hydroxyl radical and ABTS radical were evaluated with the method of Li et al. (2020). Briefly, the 0.1 mL DPPH ethanol solution (0.2 mmol/L) was added to 0.1 mL different concentrations of polyphenols solution (0–0.2 mg/mL) and the mixed solution was cultured at 37 °C for 30 min, the absorbance was measured at 517 nm. For hydroxyl radical scavenging assay. The 0.10 mL different concentrations of polyphenols solution (0–1.0 mg/mL) were successively mixed with 0.10 mL 9 mmol/L salicylic acid, 9 mmol/L FeSO4 and 0.01% H2O2. The mixture was incubated at 37 °C for 30 min and the absorbance was measured at 520 nm. In addition, the 0.10 mL different concentrations of polyphenols solution (0–0.1 mg/mL) was added to 0.15 mL ABTS radical and incubated at room temperature for 6 min for ABTS radical assay. The absorbance was measured at 734 nm. Anti-lipid peroxidation capacity was measured according to Cheng et al. (2019). Briefly, 0.10 mL different concentrations of polyphenols solution (1.00–5.00 mg/mL) were successively mixed with 0.10 mL lecithin concentrations (10 mg/mL), 0.10 mL FeSO4 (25 mmol/L) and 0.50 mL PBS (0.10 mol/L, pH 7.4), the mixture was incubated at 37 °C for 1 h, then added 0.5 mL 10% trichloroacetic acid and 0.5 mL 0.8% thiobarbituric acid, the mixture was boiled for 10 min and cooled to room temperature. The absorbance was measured at 532 nm.
2.7 Nematodes stress assay
The synchronized worms were prepared by allowing hermaphrodites to lay eggs 4 h at 20 °C (N2) or 16 °C (CL4176) and then removed the adult nematodes. Eggs were incubated on NGM plates seeded with E. coli OP50 and cultivated to L1 stage. Afterwards, 20 nematodes in L1 stage were placed in plates with or without ASP (0.8 and 1 mg/mL) and cultivated to L4 stage. Heat stress was at 35 °C, and the dead of worms was recorded at 2 h interval until all the worms died. For oxidative stress, the worms were treated with 50 μmol/L juglone in 96-well plates and incubation at 20 °C for 16 h.
2.8 Enzyme activity and MDA content assays in nematodes
The N2 worms (L4 stage) were transferred to 35 °C for 3 h and collected and washed with M9 buffer (0.3 g KH2PO4, 1.25 g Na2HPO4·12H2O, 0.5 g NaCl to 100 mL distilled water), then broken with an ultrasonic crusher (300 W). After centrifugation at 12,000 g for 10 min at 4 °C and ollected the supernatant. MDA and the activity of antioxidant enzymes (SOD, CAT) were evaluated complying with the instruction of the kits.
2.9 Paralysis assay
The paralysis assay was done according to Ma et. al. (2017). The C. elegans CL4176 in L1 stage was placed in fresh NGM plates with or without 1.0 mg/mL ASP and cultured at 16 °C for 36 h, and then, worms were moved to 25 °C to promote the production of Aβ peptide and induced paralysis. The paralyzed worms were numbered every 3 h until all nematodes paralyzed.
2.10 Aβ accumulation assay
The CL4176 transgenic nematodes were collected and fixed with 4% paraformaldehyde (pH 7.4) at 4 °C for 24 h, and then treated with permeable solution (5% β-mercaptoethanol, 1% Triton X-100, 125 mmol/L Tris, pH 7.4) for 24 h at 37 °C. Afterwards, the nematodes stained with 0.125% thioflavin T for 2 min, and distained with 50% ethanol. The fluorescence images were taken with a fluorescence microscope (Nikon DS-Ri1, Japan)). The number of Thioflavin T reactive deposits in the anterior of pharyngeal bulb was scored.
2.11 Determination of ROS level in worms
Th CL4176 transgenic nematodes were collected and washed with M9 buffer. Afterwards, the 50 nematodes were moved to a microtiter plate with M9 buffer and 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA), and incubated for 60 min at 37 °C. The fluorescence intensity at 535 nm was recorded with the excitation at 485 nm (Wan et al., 2021).
2.12 Gene expression assay
The CL4176 transgenic nematodes were collected and washed with M9 buffer. The total RNA was extracted using the acid-phenol (TRIzol) method and converted to cDNA using PrimeScript™ RT reagent kit following the prescribed protocol. The quantitative PCR was performed in triplicate with SYBR Premix Ex TaqTM II (Takara Biotechnology Co., Ltd.) and CFX96 Real-Time PCR Detection Systems (Bio-Rad, Chicago, USA). The primers used for PCR was shown in Table S1. The gene expression was normalized using the Actin as a housekeeping gene. The fold change in mRNA was calculated by 2-ΔΔCt method.
2.13 Statistical analysis
Values were means ± SDs (standard deviations) from three separate operations. All data were analyzed with SPSS 25.0, and one-way ANOVA was employed to statistical analysis. p < 0.05 was regarded as significance.
3 Results
3.1 Effect of single factor on the ASP yield
With the increase of ultrasonic power (150 W, 180 W, 210 W, 240 W, 270 W), the yield of polyphenols gradually increased first, and then decreased when the ultrasonic power higher than 240 W (Fig. 1(A)). Ultrasonic improved the extraction of polyphenols, but the damage of polyphenols structure leads to the decrease of the yield when the ultrasonic power too high. Thus, ultrasonic power from 210 W to 270 W was used for the further experiments.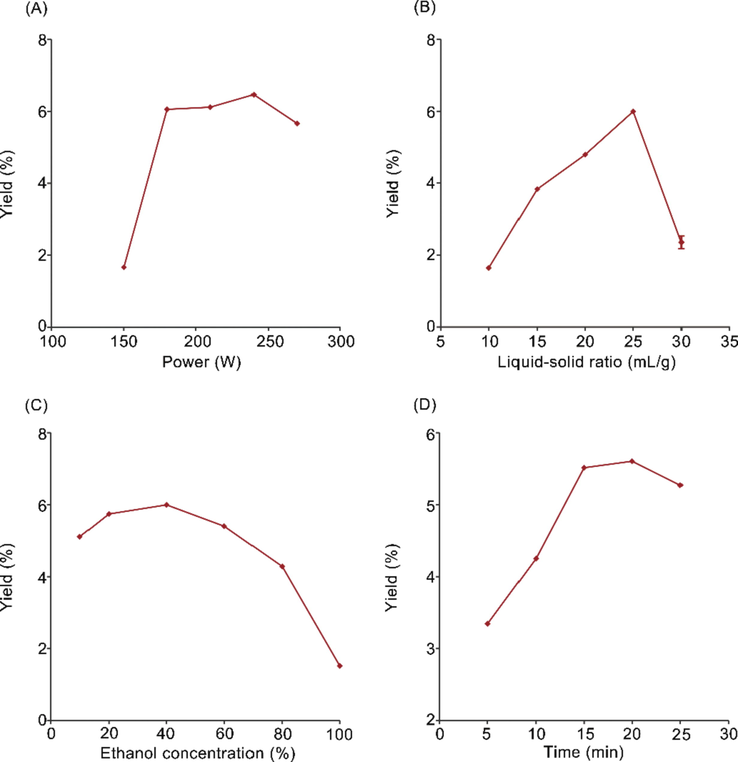
Effect of singer factor on polyphenols yield. (A) the power of ultrasonic (W), (B) the ratio of liquid–solid (mL/g), (C) the concentration of ethanol (%), (D) time (min).
The polyphenols yield was improved first with liquid–solid ratio, and then decreased when more than 25:1 (Fig. 1(B)). The liquid–solid ratio ranges from 20:1 to 30:1 (mL/g) was selected to the further experiments.
With the increase of ethanol concentration (10%, 20%, 40%, 60%, 80%, 100%), the polyphenols yield displayed a trend with first rising and then declining (Fig. 1(C)). Polyphenols reached the highest yield when ethanol concentration was 40%. Therefore, the ethanol concentration from 20% to 60% was employed for further experiments.
The yield of polyphenols reached to the maximum (5.61%) at 20 min (Fig. 1(D)). When the extraction time exceeded 20 min, the yield declined. Therefore, 15 to 25 min was used for further experiments.
3.2 Model fitting and AVOVA
As shown in Table 1, the yield of polyphenols varied from 4.30% to 8.90% among the 29 groups of tests designed by BBD. The predicted model about response variables and tests variables could be expressed by a quadratic polynomial equation as follows:
Y = 8.84 + 0.07 A1-0.15 A2-0.075 A3-0.70 A4 + 0.18 A1A2 + 0.38 A1A3-1.26 A1A4-0.06 A2A3 + 0.095 A2A4-0.31 A3A4-1.01 A12-2.03 A22-1.22 A32-1.66 A42.
Where Y represents extraction yield, A1 means ultrasonic power, A2 means liquid–solid ratio, A3 means ethanol concentration, and A4 means extraction time.
The significance of model coefficients was evaluated with t-test and P-value (Table 2). The result (P < 0.0001) indicated the model was significant. The lack of fit (F = 0.9155 and P = 0.5886) was no significant. In addition, the determination coefficient R2 implied 99.90% of the variations fitted this model. The adjusted determination coefficient (R2adj) was 0.9980, suggesting this model was significant. In addition, linear coefficient (A1, A2, A3 and A4), the quadratic coefficients (A12, A22, A32 and A42), and the interaction coefficients (A1A2, A1A3, A1A4, A2A4 and A3A4) were very significant, implying these factors significantly affected on ASP yield. A1 means ultrasonic power, A2 means liquid–solid ratio, A3 means ethanol concentration, and A4 means extraction time.
Source
Quadratic Sum
Degree of Freedom
Mean Square
F-Value
P-value; Prob > F
Model
55.252
14
3.947
983.622
< 0.0001
significant
A1
0.060
1
0.060
15.006
0.0017
A2
0.279
1
0.279
69.556
< 0.0001
A3
0.068
1
0.068
16.823
0.0011
A4
5.936
1
5.936
1479.498
< 0.0001
A1A2
0.133
1
0.133
33.204
< 0.0001
A1A3
0.562
1
0.563
140.195
< 0.0001
A1A4
6.400
1
6.400
1595.335
< 0.0001
A2A3
0.014
1
0.014
3.589
0.0790
A2A4
0.036
1
0.036
8.997
0.0096
A3A4
0.384
1
0.384
95.806
< 0.0001
A12
6.680
1
6.680
1664.983
< 0.0001
A22
26.792
1
26.792
6677.442
< 0.0001
A32
9.671
1
9.672
2410.521
< 0.0001
A42
17.951
1
17.951
4474.138
< 0.0001
Residual
0.056
14
0.004
Lack of Fit
0.0390
10
0.0039
0.9155
0.5886
not significant
Pure Error
0.0171
4
0.0043
Cor Total
55.3080
28
R2
0.9990
R2adj
0.9980
3.3 Optimization for the extraction parameters with response surface
The three-dimensional response surface plots and contour plots for the model were produced according to the regression equation to evaluate the interactions of these any two variables (Fig. 2). The polyphenols yield (Y) increased with extraction variables, and it gradually declined after the yield reached the maximum. Among the four independent variables, extraction time showed the greatest influence on polyphenols yield, following by the ratio of liquid–solid, the concentration of ethanol, and the power of ultrasonic, in that order. According to the model, the optimal conditions for ultrasonic assisted extraction of ASP were 240.12 W (the power of ultrasonic), 25.01 (the ratio of liquid–solid, mL/g), 40.14% (the concentration of ethanol) and 20.03 min (extraction time). The theoretical yield of ASP was 8.83% under these conditions.
Response surface plots for interaction of any two variables on ASP yield. (A) the power of ultrasonic and the ratio of liquid–solid, (B) the power of ultrasonic and the concentration of ethanol, (C) the power of ultrasonic and time, (D) the ratio of liquid–solid and the concentration of ethanol, (E) the ratio of liquid–solid and time, (F) the concentration of ethanol and time.
3.4 Verification of predictive model
For the convenience of operation, the extraction parameters were adjusted to extraction time 20 min, ultrasonic power 240 W, ethanol concentration 40%, and liquid–solid ratio 25 (mL/g). The experimental yield of ASP was 8.80 ± 0.04% (n = 3) under these optimal conditions, which are in good agreement with the predicted yield.
3.5 The composition of ASP
The crude polyphenols was separated by AB-8 macroporous resin and elution with 70% ethanol. The major fraction was collected and still named ASP for the following experiment. After purification, the polyphenols purity was increased from 17.34 ± 0.05% to 47.00 ± 0.04%.
The HPLC assay was performed to determine the component in ASP. The chromatogram of phenolic standards and ASP were shown in Fig. 3. Chlorogenic acid, isoorientin, vitexin, quercitrin and apiin were observed in ASP. Isoorientin and vitexin were the main compound in ASP, followed by chlorogenic acid, quercitrin, and apiin. Isoorientin and vitexin are natural C-glucosyl flavone with the robust antioxidant, neuroprotection and anti-inflammatory properties (Ziqubu et al., 2020; He et al., 2016), The results revealed the flavonoids was the main component of ASP.
The composition of ASP. (A) phenolic standards, (B) ASP.
3.6 Antioxidant activity of ASP in vitro
The scavenging activity of DPPH radical by ASP increased with its concentration (Fig. 4(A)). The scavenging rate of ASP reached 83.31% at 0.20 mg/mL, and its scavenging activity was almost as good as Vc (85.20%). The IC50 of ASP was 0.06 mg/mL. The scavenging ability of ASP to hydroxyl radical is dose-dependent (Fig. 4(B)). The scavenging rate reached the maximum (75.48%) at 1.00 mg/mL of ASP, and its IC50 was 0.72 mg/mL. ABTS was a non-physiological free radical that activated the chain reaction of free radical and destroyed the protein, DNA and lipid (Pu et al., 2019). ABTS radical scavenging capacity of ASP was positively related to its concentration (Fig. 4(C)). The ABTS radical scavenging rate of ASP achieved 86.97% at 0.10 mg/mL, and its IC50 value was 0.05 mg/mL. The anti-lipid-peroxidation capacity of ASP rose from 25.28% to 86.63% when its content increased from 1 to 5 mg/mL. Its IC50 was 2.03 mg/mL (Fig. 4(D)). These results suggested that ASP displayed favorable antioxidant activity in vitro.
Antioxidant activity of ASP in vitro. (A) the scavenging ability to DPPH radical, (B) the scavenging ability to hydroxyl radical, (C) the scavenging ability to ABTS radical, (D) anti-lipid peroxidation capacity. Each point represent mean ± SD of three independent experiments.
3.7 ASP improved the tolerance of C. elegans N2 to heat stress
Heat stress disrupted the homeostasis of oxidation/antioxidant system and caused oxidative damage in body (Habashy et al., 2019; Yang et al., 2010). The treatment of 0.8 mg/mL and 1 mg/mL ASP improved the survival time of nematodes by 4 h and 7 h compared with the control (Fig. 5(A)) under heat stress. The PT50 (50% of the nematodes died) of worms increased by 4 h after treating with 0.8 mg/mL ASP, while it increased by 5.5 h for treating with 1 mg/mL ASP compared with the control. Therefore, we chose 1 mg/mL ASP for further study.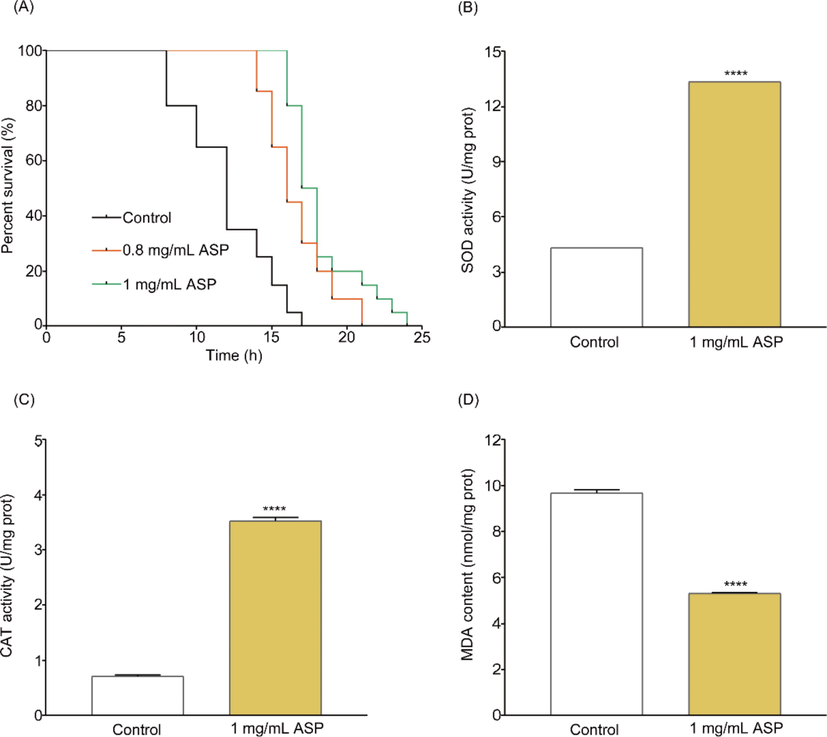
Activity of ASP under heat stress in N2. (A) the survival curves of C. elegans N2 after treatment with 1 mg/mL ASP at 35 °C, (B) the activity of SOD, (C) CAT and (D) the content of MDA of C. elegans N2 after treatment with 1 mg/mL ASP at 35 °C for 3 h. Each value is the average ± SD. **** p < 0.0001 compared with the control group.
Heat stress caused oxidative damage, and it could be restored by improving the antioxidant system, which played a key part in maintaining cellular normal metabolism (Yang et al., 2010). SOD and CAT are key antioxidant enzymes in cell. SOD can convert harmful superoxide free radicals into oxygen and H2O2, and the latter was decomposed into harmless water by CAT, thus maintaining the dynamic balance of the oxidation/antioxidant system in the organism to prevent the body from oxidation damage (Lewandowski et al., 2018). SOD and CAT activities were significantly improved after treating with 1 mg/mL ASP (Fig. 5(B), (C)). Moreover, MDA is characteristic product of lipid peroxidation, which can bind to DNA and protein lead to tissue damage, and cause cytotoxicity (Rio et al., 2005; Ho et al., 2013). After treating with ASP, the level of MDA remarkably decreased (Fig. 5(D)). These results indicated that ASP could improve the capacities of antioxidant enzymes and relieving the level of lipid peroxidation in C. elegans N2 under heat stress.
3.8 ASP relieved the paralysis and reduced Aβ deposition in C. elegans CL4176
The heat stress and oxidative stress were used to evaluate the protection effect of ASP from the oxidative damage in AD model of C. elegans. The survival time of the nematodes treated with 1 mg/mL ASP increased by 5 h, and the PT50 is up to 13 h under heat stress (Fig. 6(A)). In addition, the average survival rate of the nematodes treated with 1 mg/mL ASP increased by 38.89% under oxidative stress (Fig. 6(B)).
ASP treatment significantly reduces paralysis and Aβ plaques in C. elegans CL4176. (A) survival curves in C. elegans CL4176 treated with 1 mg/mL ASP at 35 °C, (B), survival rate in C. elegans CL4176 treated with 1 mg/mL ASP exposed to 50 μmol/L juglone for 16 h, (C) the effect of ASP on the paralysis of C. elegans CL4176 at 25 °C, (D) the fluorescence images of Aβ deposition in C. elegans CL4176 stained with thioflavin T, the white arrows pointing to Aβ deposits (E) The quantity is expressed as mean number of Aβ deposits in the head region/anterior area of the worm. Each value is the average ± SD. * p < 0.05, **** p < 0.0001 compared with the control group.
Expression of the Aβ gene in muscle of C. elegans CL4176 is associated with age-progressive paralysis, and it is widely used as AD models for evaluating the toxicity of Aβ to screening potential drugs. Therefore, we investigated the paralysis of C. elegans CL4176 treated with 1 mg/mL ASP after incubation for 46 h at 25 °C. The paralysis rate of control group was 89.32%, while it was only 40.74 ± 3.21% for the ASP treated group (Fig. 6(C)). The result indicated that the ASP protected C. elegans CL4176 from the Aβ-induced paralysis. Next, we measured the Aβ deposition in the AD model of C. elegans CL4176, we observed ASP significant decrease the number of Aβ deposition in the head area of C. elegans CL4176 compared with the control group. ASP treatment was significantly decreased by the Aβ deposition in C. elegans CL4176 (Fig. 6(D), (E)).
3.9 ASP reduced oxidative stress through P13K/AKT signal pathway in C. elegans CL4176
The accumulation of Aβ will increase the level of ROS and causing oxidative stress (Pratico, 2008). Therefore, we investigated whether the accumulation of Aβ-induced paralysis was related to the levels of ROS in C. elegans CL4176. The level of ROS was obviously decreased by ASP (Fig. 7(A)), which preliminary indicated that ASP relieved Aβ-induced paralysis probably by reducing the level of ROS.
ASP treatment reduced oxidative damage through P13K/AKT signal pathway in C. elegans CL4176, (A) the effect of ROS level in C. elegans CL4176 after treated with ASP at 25 °C for 36 h, the mRNA expression levels of skn-1 (B) akt-1, pdk-1 (C) in C. elegans CL4176 after treated with ASP at 25 °C for 36 h. Each value is the average ± SD. * p < 0.05, ** p < 0.01, **** p < 0.0001 compared with vehicle control group.
The transcription factor skinhead-1 (SKN-1) was involved in oxidative stress (Li et al., 2019). Besides, oxidative stress can damage the P13K/AKT signaling pathway that causes neuron damage and AD (Jiang et al., 2016). AD can be alleviated by the P13K/AKT signaling pathway (Sun et al., 2020). ASP improved the expression of skn-1 and down-regulated the transcription of akt-1 and pdk-1 (Fig. 7(B), (C)). These results revealed that ASP might alleviate AD by down-regulation of P13K/AKT signaling pathway.
4 Discussion and conclusion
Polyphenols as a type of secondary metabolic product have been drawn more and more attentions. UAE is considered as a green method for polyphenols extraction from the natural material. Compared with the traditional extraction techniques, ultrasonic assisted extraction saved energy, time, and solvents (Ameer et al., 2017). We optimized the process of ultrasonic assisted extraction polyphenols from the leaves of A. spinulosa and the extraction yield was 8.80 ± 0.04%. Detti et al. (2020) extracted the polyphenols from Pistacia lentiscus L. leaves with UAE, the yield of polyphenols was 5.13 ± 0.18%. In addition, Chmelova et al. (2020) found that the polyphenols from Picea abies bark with UAE was higher than solvent extraction.
The antioxidant activity of polyphenols is responsible for protecting cells. There are two main mechanisms for the antioxidant activity of polyphenols. Firstly, the functional group of phenol can provide free radical R• with hydrogen atom. In this case, the polyphenols act as chainbreaker. Secondly, polyphenols form a radical cation through transferring a single electron to the free radical (Quideau et al., 2011). In addition, polyphenols can reduce the cell oxidative damage through the antioxidant enzyme system. We observed that ASP had good scavenging abilities to DPPH radical, hydroxyl radical and ABTS radical and good anti-lipid-peroxidation capacity in vitro. Further, ASP can confer C. elegans N2 with higher tolerance to heat stress, improved the capacities of SOD and CAT, and reduced the content of MDA. The polyphenols of apple leaves had good scavenging capacity of DPPH radical, and reduced the H2O2-induced oxidant damage in rat hippocampal neurons by improved SOD and lactate dehydrogenase (LDH) activities (Lu et al., 2019). Vitexin relieved oxidative damage by decreasing the content of MDA, enhancing the activity of SOD and CAT (Li et al., 2021).
Abnormal Aβ accumulation is one of the main etiologies in AD (Ashrafian et al., 2020). There is evidence suggesting that Aβ accumulation will trigger neurotoxicity and oxidation (Shankar et al., 2008). The antioxidant could inhibit the aggregation of Aβ and contribute to preventing the occurrence or progression of AD (Petrovic et al., 2020). 1 mg/mL ASP significantly relieved the Aβ-induced paralysis and reduced the Aβ plaques in C. elegans CL4176. The transcription factor SKN-1 was orthologous gene with Nrf-2 (NF-E2-related factor 2) of human, and it was crucial in defending against oxidative damage in C. elegans (Li et al., 2017). Reduction of Aβ-induced toxicity through lessening oxidative damage in AD patients is important for mitigating the AD process. In present studies, we observed ASP decreased ROS level and enhanced the expression of SKN-1 in AD model of C. elegans CL4176. In addition, ASP down-regulated the expression of pdk-1 and akt-1 in P13K/AKT signaling pathway in C. elegans CL4176. Our results indicated that ASP might alleviate the Aβ-induced oxidative damage through the P13K/AKT signaling pathway, thus relieving the paralysis symptom of C. elegans CL4176. In H2O2-treated APPswe cells, rutin and quercetin attenuated and reversed Aβ25–35 fibril formation and improved intracellular GSH level and maintained the redox homeostasis (Jimenez-Aliaga et al., 2011). In addition, flavone, apigenin, luteolin, kaempferol and quercetin reduced the level of ROS which was induced by Aβ1-42 oligomer (Pate et al., 2017).
In summary, the polyphenols was extracted from the leaves of A. spinulosa (ASP) with ultrasonic-assisted extraction. The extract yield of ASP reached 8.80 ± 0.04% under 240 W of ultrasonic power, 40% of ethanol concentration, 25 (mL/g) of liquid–solid ratio and 20 min of extraction time. ASP was composed of Chlorogenic acid, isoorientin, vitexin, quercitrin and apiin. ASP displayed strong antioxidant activity in vitro, and it can alleviate Aβ-induced toxicity and oxidative damage through down-regulated of P13K/AKT signaling pathway in AD model of C. elegans CL4176. ASP might contribute to treat or prevent the AD and other neurodegenerative disease in humans. However, our present study only preliminarily demonstrated the bioactivities of ASP, but the relationship between the bioactivities and structure of ASP is unclear. Therefore, we should research the structure and explore the direct target and detail regulation pathway network of ASP for AD in further work, and analyze the conformational relationship between structure and bioactivities.
Acknowledgments
This work was supported by the Sichuan Agricultural University.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Natural polyphenols: An overview. Int. J. Food Prop.. 2016;20:1689-1699.
- [CrossRef] [Google Scholar]
- Green extraction methods for polyphenols from plant matrices and their byproducts: A review. Compr. Rev. Food Sci. Food Saf.. 2017;16:295-315.
- [CrossRef] [Google Scholar]
- Review on Alzheimer's disease: inhibition of amyloid beta and tau tangle formation. Int. J. Biol. Macromol.. 2020;167:382-394.
- [CrossRef] [Google Scholar]
- Effect of peach puree incorportion on cookie quality and on simulated digestion of polyphenols and antioxidant properties. Food Chem.. 2020;333:127464
- [CrossRef] [Google Scholar]
- Chemical constituents in the leaves of Alsophila spinulosa. Acta Bot Boreal-Occident Sin.. 2008;28:1246-1249.
- [Google Scholar]
- The antioxidant activities of carboxymethylated garlic polysaccharide and its derivatives. Int. J. Biol. Macromol.. 2019;140:1054-1063.
- [CrossRef] [Google Scholar]
- Ultrasonic-assisted extraction of polyphenols and antioxidants from Picea abies bark. J. Biotechnol.. 2020;314:25-33.
- [CrossRef] [Google Scholar]
- Optimization of a green ultrasound-assisted extraction of different polyphenols from Pistacia lentiscus L. leaves using a response surface methodology. Plants (Basel). 2020;9:1482.
- [CrossRef] [Google Scholar]
- Natural polyphenols effects on protein aggregates in Alzheimer's and Parkinson's prion-like diseases. Neural Regen Res.. 2018;13:955-961.
- [CrossRef] [Google Scholar]
- Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol.. 2019;63:1569-1584.
- [CrossRef] [Google Scholar]
- A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia. 2016;115:74-85.
- [CrossRef] [Google Scholar]
- Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol.. 2013;1:483-491.
- [CrossRef] [Google Scholar]
- Oxidative stress: a major pathogenesis and potential therapeutic target of antioxidative agents in Parkinson's disease and Alzheimer's disease. Prog. Neurobiol.. 2016;147:1-19.
- [CrossRef] [Google Scholar]
- Quercetin and rutin exhibit antiamyloidogenic and fibril-disaggregating effects in vitro and potent antioxidant activity in APPswe cells. Life Sci.. 2011;89:939-945.
- [CrossRef] [Google Scholar]
- Inhibition of copper-zinc superoxide dismutase activity by selected environmental xenobiotics. Environ. Toxicol. Pharmacol.. 2018;58:105-113.
- [CrossRef] [Google Scholar]
- O-GlcNAcylation of SKN-1 modulates the lifespan and oxidative stress resistance in Caenorhabditis elegans. Sci. Rep.. 2017;7:43601.
- [CrossRef] [Google Scholar]
- Arginine methylation of SKN-1 promotes oxidative stress resistance in Caenorhabditis elegans. Redox Biol.. 2019;21:101111
- [CrossRef] [Google Scholar]
- Physicochemical properties, antioxidant and antiproliferative activities of polysaccharides from Morinda citrifolia L. (Noni) based on different extraction methods. Int. J. Biol. Macromol.. 2020;150:114-121.
- [CrossRef] [Google Scholar]
- Vitexin alleviates high-fat diet induced brain oxidative stress and inflammation via anti-oxidant, anti-inflammatory and gut microbiota modulating properties. Free Radical Bio Med.. 2021;171:332-344.
- [CrossRef] [Google Scholar]
- Preparative purification of polyphenols from sweet potato (Ipomoea batatas L.) leaves by AB-8 macroporous resins. Food Chem.. 2015;172:166-174.
- [CrossRef] [Google Scholar]
- Comprehensive evaluation of effective polyphenols in apple leaves and their combinatory antioxidant and neuroprotective activities. Ind Crop Prod.. 2019;129:242-252.
- [CrossRef] [Google Scholar]
- Peptides from sesame cake reduce oxidative stress and amyloid-β-induced toxicity by upregulation of SKN-1 in a transgenic Caenorhabditis elegans model of Alzheimer’s disease. J. Funct. Foods. 2017;39:287-298.
- [CrossRef] [Google Scholar]
- Society the classification of the Cyatheaceae by R. M. Tryon. Am Fern J.. 1971;61:142-143.
- [CrossRef] [Google Scholar]
- Anthoxanthin polyphenols attenuate abeta oligomer-induced neuronal responses associated with Alzheimer's disease. CNS Neurosci. Ther.. 2017;23:135-144.
- [CrossRef] [Google Scholar]
- Lipid peroxidation and antioxidant supplementation in neurodegenerative diseases: A review of human studies. Antioxidants.. 2020;9:1128.
- [CrossRef] [Google Scholar]
- Oxidative stress hypothesis in Alzheimer's disease: a reappraisal. Trends Pharmacol. Sci.. 2008;29:609-615.
- [CrossRef] [Google Scholar]
- An in vitro comparison of the antioxidant activities of chitosan and green synthesized gold nanoparticles. Carbohydr. Polym.. 2019;211:161-172.
- [CrossRef] [Google Scholar]
- Plant polyphenols: chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. Engl.. 2011;50:586-621.
- [CrossRef] [Google Scholar]
- A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr Metab Cardiovasc Dis.. 2005;15:316-328.
- [CrossRef] [Google Scholar]
- Optimization of ultrasound-assisted extraction of polyphenols from wheatgrass (Triticum aestivum L.) J. Food Sci. Technol.. 2020;57:2809-2818.
- [CrossRef] [Google Scholar]
- Amyloid-beta protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory. Nat. Med.. 2008;14:837-842.
- [CrossRef] [Google Scholar]
- GABAB receptor-mediated PI3K/Akt signaling pathway alleviates oxidative stress and neuronal cell injury in a rat model of Alzheimer's disease. J. Alzheimers Dis.. 2020;76:1513-1526.
- [CrossRef] [Google Scholar]
- Physicochemical characterization and antioxidant effects of green microalga Chlorella pyrenoidosa polysaccharide by regulation of microRNAs and gut microbiota in Caenorhabditis elegans. Int. J. Biol. Macromol.. 2021;168:152-162.
- [CrossRef] [Google Scholar]
- Effects of acute heat stress and subsequent stress removal on function of hepatic mitochondrial respiration, ROS production and lipid peroxidation in broiler chickens. Comp Biochem Phys C.. 2010;151:204-208.
- [CrossRef] [Google Scholar]
- Alleviation of symptoms of Alzheimer's disease by diminishing Abeta neurotoxicity and neuroinflammation. Chem. Sci.. 2019;10:10149-10158.
- [CrossRef] [Google Scholar]
- The role of 6-gingerol on inhibiting amyloid beta protein-induced apoptosis in PC12 cells. Rejuv Res.. 2015;18:413-421.
- [CrossRef] [Google Scholar]
- Isoorientin: A dietary flavone with the potential to ameliorate diverse metabolic complications. Pharmacol. Res.. 2020;158:104867
- [CrossRef] [Google Scholar]
Appendix A
Supplementary material
Supplementary data to this article can be found online at https://doi.org/10.1016/j.arabjc.2022.103707.
Appendix A
Supplementary material
The following are the Supplementary data to this article:Supplementary data 1
Supplementary data 1




